The discovery of rapid blight is the story of a new plant disease whose unusual biology, inconspicuous morphology and apparently low economic impact nearly consigned it to oblivion. It is also a story of what can be so rewarding about plant pathology — the excitement of discovery, the appeal of unraveling the complex interactions among plant, pathogen and environment, and the joy of collaboration. For it was only through a cooperative effort among a privately funded researcher, publicly funded university and USDA researchers, and the golf course industry that the identity of a new terrestrial pathogen was discovered, its economic impact better understood and strategies for its management developed.
Early Observations of a New Disease
The story first begins during the winter of 1995 when David Zahrte, the superintendent of Santa Ana Country Club in Southern California submitted a sample to the Pace diagnostic lab, located in San Diego, California. Zahrte is a highly experienced manager of 19 Poa annua (annual bluegrass) putting greens. His worried description of greens that had seemed fine on Friday afternoon but were suddenly covered with mysterious looking patches of dead turf on Monday was a cause for concern (Fig 1.).
The foliage of the affected turf was yellow and brown and had a water-soaked appearance, while the roots of the plant seemed unaffected. But, microscopic observation of the sample revealed none of the usual winter time Poa diseases, such as Fusarium patch. There were no obvious signs of fungal pathogen invasion such as mycelia or spores. No insect or nematode pests or their damage were observed, whereas nutritional and cultural problems such as anaerobic soils or nutrient imbalances were also ruled out (29). The only detected abnormality was the presence of many thin walled, spindle-shaped cells, measuring about 6 × 16 µm, packed inside the foliage mesophyll cells (Fig. 2) (28)
 |
|
 |
Fig. 1. Initial observation of a mysterious disease (later to be named rapid blight) on an annual
bluegrass putting green at Santa Ana Country Club, Ca in 1995 (left) and in 1997 (right) illustrating the range
of field symptoms from small patches to larger coalescing areas on annual bluegrass (photo, L. J. Stowell).
| |
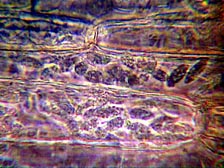
Fig. 2. Spindle-shaped cells of the rapid blight pathogen in leaf tissue. Spindle cells are about 6 µm wide and 16 µm long (photo, L. J. Stowell). |
|
Were it not for the sheer number of the spindle-shaped cells, they could easily have been mistaken for cellular organelles belonging to the plant. In fact, one mycologist suggested the spindle cells of the suspected pathogen might be just that, plant chloroplasts. However, when turfgrass samples were maintained in the laboratory in a moist chamber, the disease spread from diseased to healthy plants. There was a constant association between the presence of the spindle cells and diseased plant tissue. Unfortunately this organism did not resemble any other type of documented turfgrass pathogen.
A Series of Dead Ends
Early attempts to identify the organism causing this new disease were frustrating. Identification through standard taxonomic keys and attempted isolation on standard culture media were fruitless. Initially the organism was tentatively identified based on the morphology of its cells as a chytridiomycete. But attempts by Jim Adaskaveg of the University of California, Riverside, to isolate a chytridiomycete on specialized media from infected samples were unsuccessful. Mycologists suggested the organism might instead be a protozoan, a single-celled animal. Protozoologists countered suggesting the organism was most likely a chytridiomycete. Both were mistaken. In the end it would be discovered that not only was this organism a unique pathogen to turfgrasses, but it was the first observation of this type of organism attacking land plants of any kind.
Early Progress on Disease Control: Putting Out the Fire
From 1995 through 1998, an increasing number of samples of infected annual bluegrass began to arrive at Pace’s diagnostic laboratory from locations throughout California, some samples of annual bluegrass arriving from Colorado, and additional samples of rough bluegrass (Poa trivialis), used for overseeded bermudagrass putting greens, arriving from Nevada. At first glance, the problem appeared to be spreading, but it is equally likely that word had begun to travel about a new and mysterious disease, and superintendents were anxious to see if their golf course turf was being damaged by the disease. Control methods for the disease were needed even though the causal organism had not yet been identified.
The late Houston Couch of Virginia Tech was contacted for input on potential control strategies. Based on Pace’s descriptions of the thin-walled, spindle-shaped cells, Couch suggested that mancozeb might be an effective control agent due to its mode of action as a general membrane disruptor. His guess was indeed correct, and shortly afterwards, mancozeb became the first recognized treatment for prevention and to limit spread of the disease. This material was used under FIFRA Section (2ee), which permits the use of a registered pesticide on a pest that does not appear on the label, as long as the product is used on a labeled crop and all use and handling conditions on the label are followed.
The Project Gains Steam and Collaborators
Initially there appeared to be little interest in this new turfgrass disease in the academic world or the agrichemical industry. However, as the disease was identified in additional hosts and from additional locations, interest slowly grew. In 1999, four years after the disease was first described from California Poa annua greens, superintendent Mick Twito of Estrella Mountain Ranch in Arizona submitted samples from a third host, perennial ryegrass (Lolium perenne). And in December of 2000, the first sample from the eastern U.S. was diagnosed when superintendent Tommy Witt of Cassique Golf Course in South Carolina, and then-President of the Golf Course Superintendent’s Association of America submitted a sample of rough bluegrass that contained the same spindle-shaped cells in diseased foliage.
| |
 |
|
Fig. 3. Comparison of symptoms of a new disease (later to be known as rapid blight) on rough
bluegrass in South Carolina (top) (photo, S. B. Martin), ryegrass in Arizona (bottom left)
(photo, L. J. Stowell), and bentgrass in California (bottom right) (photo, L. J. Stowell).
The occurrence of the disease in South Carolina was important because it brought Samuel "Bruce" Martin, of Clemson University, into the project. Martin’s lab took rapid action in 2001 by initiating a series of chemical management, host range, cultural, biological, and molecular studies. Steven Alderman, a USDA forage pathologist at Oregon State University also became involved, investigating the potential for infection and transmission within the perennial ryegrass and rough bluegrass production seed fields in the Pacific Northwest. Fortunately he ruled out seed-borne transmission as the cause of the initial disease outbreaks.
To further support the productive collaboration and information exchange that was emerging among superintendents and turf researchers, a working group was formed in 2001. The group consisted of superintendents from 60 golf courses around the country that were affected by this disease
Martin and Stowell were uncomfortable with the fact that golf course superintendents and others were using the name ‘chytrid’ to describe this unknown disease. It was suspected to be caused by some unknown organism resembling those in the Chytridiomycota, but it had not been confirmed. So, in telephone conversations, Martin and Stowell jointly coined the name rapid blight, which adequately described the consequences of the disease when it occurred in epidemic proportions (14,15,16).
The working group’s first action was to provide funds to support Martin’s initial research on disease control. Although the funding was insufficient to support a full fledged project, the United States Golf Association (USGA) soon provided a substantial grant to Martin’s group to support further research. At the same time, Kurt Desiderio, a Pace Working Group member from Saticoy Country Club in 2001, was growing frustrated by the incomplete control of the disease with mancozeb. Based on a quick screening trial that he conducted on the golf course, he found that trifloxystrobin (Compass) was effective at stopping the disease. This was a surprise, since repeated tests with the closely related azoxystrobin (Heritage) had yielded no positive results. But Martin immediately followed up by placing Compass and pyraclostrobin (which was soon to be labeled as Insignia) in his next round of screening tests, and Desiderio’s observation was confirmed. Trifloxystrobin was the first material shown to be more effective than mancozeb in control of rapid blight. In 2003, Insignia was labeled for turf disease control, and included rapid blight on the label.
Identification of the Organism Causing Rapid Blight, Finally!
The collaboration among scientists and superintendents received another boost when Mary Olsen, University of Arizona Extension Plant Pathologist became involved amidst an increase in the number of Arizona golf courses that were suffering with the disease on greens, fairways and even roughs. Working closely with Donna Bigelow, Dave Kopec, and Robert Gilbertson, Olsen initiated laboratory and field research on rapid blight in 2002. The Pace Working Group was able to provide some funds to support this research. Olsen’s work soon led to the long-awaited identification of the organism and the naming of a new species.
In 2002, Robert L. Gilbertson, Professor Emeritus of Mycology at the University of Arizona, recognized the spindle-shaped cells in a rapid blight affected rough bluegrass sample from a golf course in central Arizona, as similar to those that cause a disease of eelgrass in marine estuaries (18). The organism causing the wasting disease of eelgrass, Labyrinthula zosterae, is a marine slime mold. On the basis of shared morphological characteristics associated with the size and shape of its spindle-shaped vegetative cells and growth characteristics, Olsen and her associates proposed that the rapid blight pathogen was a member of genus Labyrinthula.
Olsen subsequently was able to grow the cells isolated from diseased turfgrass tissue on an artificial medium developed specifically for Labyrinthula (27). Cells harvested from cultures were used to inoculate healthy Poa trivialis and perennial ryegrass. Symptoms identical to those observed in the field developed on all inoculated plants. Labyrinthula was then re-isolated from the inoculated diseased turfgrass (19). This technique of isolation, pure culture, and re-infection is known as Koch’s postulates and is considered proof that the organism is the true pathogen and cause of disease. Olsen’s group named the rapid blight causing organism Labyrinthula terrestris (3). It is the only Labyrinthula known to attack a land plant; all others occur in marine environments.
Description of Labyrinthula

Fig. 4. Vegetative cells of L. terrestris illustrating longitudinal cell division (photo, D. Bigelow). |
|
Labyrinthula has been classified in different ways since it was first described in 1867 (6). It is now placed in the kingdom Chromista (also called the Stramenopiles) with organisms such as diatoms and the Oomycetes (species of Pythium and Phytophthora are in this group), but it is not closely related to these organisms.
Labyrinthula terrestris vegetative cells are fusiform, averaging about 6 × 16 μm, and usually divide longitudinally (Fig. 4). Labyrinthula terrestris forms digitate colonies in an extracellular network produced by specialized organelles called bothrosomes and can move along these networks at a notable speed (Fig. 5). Cells contain various sized vacuoles, numerous lipid droplets, and have a central nucleus with a large nucleolus (Fig. 6). As cells multiply, colonial networks are formed and expand to as wide as 5/32 inch (4 millimeters) in 24 hours on agar culture media. After about a week the cells migrate into rounded clumped aggregates that range from 0.1 - 0.5 mm in diameter. Sori or reproductive cells have never been seen (3,20).
| |
 |
|
 |
|
| |
Fig. 5. Digitate colonies an extracellular network produced by bothrosomes (photo, D. Bigelow). |
|
Fig. 6. Transmission electron micrograph illustrating the central nucleus with a large nucleolus (photo, D. Bigelow). |
|
Given the quick emergence and increasing incidence of rapid blight disease on golf course turf, a full characterization of the pathogen through DNA studies was critical. Paul Peterson, a post-doctoral fellow in Martin’s group, collected isolates of L. terrestris from rapid blight-affected turfgrasses from the east and west coasts as part of a USGA assisted, nationwide Rapid Blight - Disease, Water and Soil Survey that Martin’s group initiated in 2003. In close cooperation with the Fungal Genomics Laboratory at N.C. State University, under the direction of Ralph Dean, the rapid blight Labyrinthula sp. were found to differ from other described and some nondescribed species of Labyrinthula, for example, L. zosterae, which causes wasting disease of eelgrass (7,8). This work confirmed and supported the original morphological characterization and identification of the rapid blight pathogen by Olsen. Additional gene sequencing to examine genetic diversity among rapid blight pathogens is currently underway (9).
Environmental Conditions Favoring Rapid Blight Occurrence
In most, but not all, cases of rapid blight disease occurrence have been associated with saline irrigation water and an accumulation of salt in the soil. To further evaluate this relationship, Martin’s group sent out a nationwide request to golf courses with suspected rapid blight to submit samples of affected turf, irrigation water and soil for analysis. Evaluation of the samples was then carried out as part of the above-mentioned Rapid Blight Disease, Water and Soil Survey, and a database on soil, water, and weather parameters associated with disease outbreaks was compiled. Interestingly, in the Carolinas, rapid blight outbreaks occurred concurrent with drought and applications of high salinity irrigation water (>2.5 dS/M or >1,600 ppm). However, on some western golf course sites, the disease may also occur at lower salinity levels, 0.5 – 1.5 dS/M or 320 - 960 ppm) based on the survey (Peterson, unpublished).
Subsequent greenhouse trials showed that little disease occurred in susceptible perennial ryegrass and Kentucky bluegrass cultivars which had received applications of irrigation water ≤1.3 dS/m (4,5). Disease severity increased with increased salinity. Results from Olsen’s group were quite similar. Plants irrigated with low salinity water (0.5 dS/m) show no symptoms of disease, but become infected. As salinity of the irrigation water increases from 0.8 dS/m to 4.0 dS/m disease severity also increases (11,12,21). Managing irrigation water to reduce salt accumulation in soils is an effective way to reduce the occurrence of rapid blight.
Geographic Distribution and Host Range
Rapid blight was initially identified on golf courses in the United States on annual bluegrass, rough bluegrass, and perennial ryegrass. Although the disease has been documented on creeping bentgrass by Martin and Stowell, it has rarely occurred. By 2005, Entwistle, in cooperation with Olsen, described the disease attacking colonial bentgrass and annual bluegrass golf course greens in the United Kingdom (10). The report from the U.K. was notable for its northern location; until then, rapid blight had been isolated only from 11 U.S. states (30).
Peterson evaluated forty-nine different cool-season turfgrass species in hopes of identifying specific turfgrass species and cultivars with tolerance to rapid blight. All the grasses examined were susceptible to rapid blight but at markedly varying levels. Mean disease severity levels among the cultivars tested ranged from <1% to >90% (25). Bentgrasses (colonial & velvet), bluegrasses (annual and rough), most ryegrasses, crested dogstail, hairgrass, and wheatgrasses were susceptible. The grass species most tolerant to rapid blight were the slender creeping red fescues, creeping bentgrasses and some alkaligrasses (Fig. 7 and Fig. 8). Similar results were obtained by Olsen’s group in Arizona (2,13,20).
 |
|
 |
|
Fig. 7. The devastating effects of rapid blight on annual bluegrass from cool-season turfgrass susceptibility/tolerance studies. Non-inoculated, left; inoculated right – 16 days post-inoculation (photo, P. D. Peterson). |
|
Fig. 8. ‘Dawson’ slender creeping red fescue, the most tolerant grass from cool-season turfgrass susceptibility/tolerance studies. Non-inoculated, left; inoculated right – 16 days post-inoculation. (photo, P. D. Peterson). |
According to Olsen, bermudagrass shows no symptoms of rapid blight, but in a survey of two golf courses where rapid blight occurred in cool-season grasses used for over-seeding, L. terrestris was isolated from bermudagrass roots and stolons during the summer after the cool-season turfgrasses had died out.
The combined observations of these experiments showed several cool-season turfgrasses to be tolerant of rapid blight under conditions of moderately high salinity stress. Some of these grasses may be suitable for overseeding where rapid blight is a chronic problem. In SC, some golf courses are using seed blends of rough bluegrass and alkaligrass with acceptable results. Potential exists for the use of certain creeping bentgrass cultivars or slender creeping red fescues for overseeding as well, although slow rates of germination and establishment may be complicating factors to consider (24,25).
The host range of L. terrestris may not be limited to turfgrass. In a preliminary study, non-salt tolerant cultivars of rice, lettuce and radish as well as salt tolerant varieties of alfalfa, barley, and wheat were screened in the greenhouse and laboratory to determine if L. terrestris could infect plants other than turfgrasses. Wheat, barley and rice plants were infected, became symptomatic and died. Radish and lettuce were infected but non-symptomatic. Alfalfa was not infected and exhibited no symptoms (1).
Other Factors Affecting Labyrinthula terrestris
A thorough knowledge of the biology and lifestyle of a disease organism is helpful in managing and controlling disease caused by that organism. For this purpose, Martin’s and Olsen’s groups conducted a series of controlled environment experiments to determine the growth characteristics of the pathogen.
In laboratory studies, Olsen found that L. terrestris grew well at 15 C to 30 C, but grew very slowly at 4°C and not at all at 40°C (20). Peterson found similar results when he evaluated the relative growth of 14 different L. terrestris. isolates collected across the United States under varying degrees of temperature and levels of salinity. Growth parameters were studied and measured on solid media. The results of these experiments from Martin’s lab indicated that L. terrestris grew best in a range between 22°C to 26°C (23).
L. terrestris isolates grew well over a relatively wide range of salinity levels from 3.5 to 10.5 dS/m (26). These results suggest that different L. terrestris isolates vary in their salt requirements and that east coast isolates may require higher salinities for optimal growth than west coast isolates.
Wounding is not necessary for L. terrestris to enter the plant, and L. terrestris moves easily from infected plants to non-infected plants when only a few leaves are touching or when plants share common drainage water (20).
Chemical and Cultural Control
Field trials to determine efficacy of selected chemicals for control of rapid blight have been conducted at several sites. The most effective chemicals for prevention of rapid blight identified so far are pyraclostrobin (Insignia), trifloxystrobin (Compass), and mancozeb (Fore, Protect). Compass or Insignia mixed or rotated with mancozeb gives good control if applied preventively, while curative applications of chemicals may contain the disease but do not eradicate it (13,14,17,21).
Cultural control requires a variety of strategies including leaching programs to reduce soil salts. Leaching alone is not sufficient in many cases, however. For this reason, the selection of rapid blight-tolerant overseeding varieties holds promise. Blending fast-establishing susceptible grasses (rough bluegrasses or moderately susceptible grasses like the perennial ryegrasses) with tolerant grasses (alkaligrasses, creeping bentgrasses, and slender creeping red fescues) may help to reduce the risk of devastating epidemics of rapid blight (22,23,24,25).
Where did Labyrinthula terrestris Come From and Where is it Going?
If the preliminary results of molecular analysis by Martin’s group continue to yield genetic sequence data that lacks variation, there may be a recent common ancestor of the rapid blight pathogens. That ancestor currently appears to be most closely related to a Labyrinthula species that attacks Spartina alterniflora, a true grass (family Poaceae) in marine environments (9). When and how the jump from marine environments to terrestrial plants was made may never be known but the search for the answer is intriguing and will occupy plant pathologists for years to come.
There are many more questions to answer concerning rapid blight and Labyrinthula as a plant pathogen. From what we know now, rapid blight affects a broad range of cool season turfgrasses that show an increased severity of symptoms as soil salinities increase. Soil salinity problems are likely to increase in the future as competition for high quality water, increased use of recycled water on golf courses, and drought conditions occur. As a result, turf managers will need to develop management strategies that cope with the potential for increased rapid blight attacks, and the possibility of infections in agricultural crops will need to be further examined. In the meantime, a successful integrated approach has been identified that relies on a combination of cultural practices and chemical control.
Literature Cited
1. Bigelow, D. M., and Olsen, M. W. 2004. Investigations of the host range of Labyrinthula terrestris, a new turfgrass pathogen. 2004. The 2004 Turfgrass, Landscape and Urban IPM Research Summary. Coll. Agric. Life Sci., Univ. of Ariz.
2. Bigelow, D. M., and Olsen, M. W. 2004. Infection of Selected Turfgrasses by Labyrinthula terrestris. The 2004 Turfgrass, Landscape and Urban IPM Research Summary. Coll. Agric. Life Sci., Univ. of Ariz.
3. Bigelow, D. M., Olsen, M. W., and Gilbertson, R. L. 2005. Labyrinthula terrestris sp. nov., a new pathogen of turfgrass. Mycologia 97:185-190
4. Camberato, J. J., Peterson, P. D., and Martin, S. B. 2005. Salinity and salinity tolerance alter rapid blight disease occurrence. (Abstr.) Phytopathology 95.
5. Camberato, J. J., Peterson, P. D., and Martin, S. B. 2005. Salinity and what it means to turf after rapid blight. Carolinas Green March-April.
6. Cienkowski, L. 1867. Ueber den Bau und die Entwicklung der Labyrinthuleen. Archiv für mikroscopische Anatomie 3:274-310.
7. Craven, K. D., Peterson, P. D., Mitchell, T. K., and Martin, S. B. 2004. Phylogenetic relationships between the rapid blight pathogen of turf and aquatic protests in the genus Labyrinthula. (Abstr.) Phytopathology 94:S22.
8. Craven, K. D., Peterson, P. D. Windham, D. E., Mitchell, T. K., and Martin, S. B. 2005. Molecular identification of the turfgrass rapid blight pathogen. Mycologia 97:160-166.
9. Craven, K. D., Peterson, P. D., Martin, S. B., and Mitchell, T. K. 2005. Genetic diversity among rapid blight pathogens of cool-season turfgrasses. Inoculum 56:14.
10. Entwistle, C. A., Olsen, M. W., and Bigelow, D. M. 2005. First report of a Labyrinthula spp. causing rapid blight of Agrostis capillaris and Poa annua on amenity turfgrass in the UK. Plant Pathol. (In press).
11. Kohout, M. J., Bigelow, D. M., and Olsen, M. W.. 2004. Effect of salinity and cutting on symptom development of rapid blight of perennial rye. (Abstr.) Phytopathology 94:S54.
12. Kohout, M. J., Bigelow, D. M., and Olsen, M. W. 2004. Effect of salinity on symptom development of rapid blight on perennial rye. The 2004 Turfgrass, Landscape and Urban IPM Research Summary. The Univ. of Arizona.
13. Kopec, D., Olsen, M. W., Gilbert, J. J., Bigelow, D. M., and Kohout, M. J. 2004. Cool-season grass response to rapid blight disease. Golf Course Management 72:78-81.
14. Martin, B., Stowell, L., and Gelernter, W. 2002. Rough bluegrass, annual bluegrass and perennial ryegrass hit by new disease. Golf Course Management April 2002: 61-65.
15. Martin, S. B., Stowell, L. J., Gelernter, W. D., and Alderman, S. C. 2002. (Abstr.) Rapid blight: A new disease of cool season turfgrasses. Phytopathology 92:S52.
16. Martin, S. B., Stowell, L. J., Gelernter, W. D., and Alderman, S. C. 2002. Rapid blight: A new disease of cool season turfgrasses. Turfgrass Program, Clemson Univ.
17. Martin, S. B., Olsen, M. W., Peterson, P. D., and Camberato, J. J. 2004. Evaluation of fungicides for control of rapid blight on cool season grasses. (Abstr.) Phytopathology 94:S66.
18. Muehlstein L. K., Porter, D., and Short, F. T. 1991. Labyrinthula zosterae sp. nov., the causative agent of wasting disease of eelgrass, Zostera marina. Mycologia 83:180-191.
19. Olsen, M. W., Bigelow, D. M., Gilbertson, R. L., Stowell, L. J., and Gelernter, W. D. 2003. First report of a Labyrinthula sp. causing rapid blight disease of rough bluegrass and perennial ryegrass. Plant Disease 87:1267.
20. Olsen, M. W., Bigelow, D. M., Kohout, M. J, Gilbert, J. J., and Kopec, D. 2004. Rapid blight: A new disease of cool-season turfgrass. Golf Course Management 72:87-91.
21. Olsen, M. W., and Gilbert, J. 2004. Evaluation of Fungicides for Control of Rapid Blight of Poa trivialis.. The 2004 Turfgrass, Landscape and Urban IPM Research Summary. Coll. Agric. Life Sci., Univ. of Ariz.
22. Peterson, P. D., Martin, S. B., and Camberato, J. J. 2004. Rapid blight disease of cool-season grasses. TGIF Record No. 97542. USGA Turfgrass and Environ. Res. Online 3:1-6.
23. Peterson, P. D., Martin, S. B., and Camberato, J. J. 2004. Determining the turfgrass host range of Labyrinthula sp., the causal agent of rapid blight. (Abstr.) Phytopathology 94:S83.
24. Peterson, P. D., Martin, S. B., and Camberato, J. J. 2004. Rapid blight disease of cool-season grasses. USGA Green Section Record 42:20-23.
25. Peterson, P. D., Martin, S. B., and Camberato, J. J. 2005. Tolerance of cool-season turfgrasses to rapid blight disease. Applied Turfgrass Science doi:10.1094/ATS-2005-0328-01-RS.
26. Peterson, P. D., Martin, S. B., Camberato, J. J., and Fraser, D. E. 2005. The effect of temperature and salinity on growth of the turfgrass pathogen, Labyrinthula terrestris. (Abstr.) Phytopathology 95.
27. Porter, D. 1987. Labyrinthulomycetes. Pages 110-113 in: Zoosporic Fungi in Teaching and Research. M. S. Fuller and A. Jaworski, eds. Athens, Georgia: Southeastern Publishing Corporation.
28. Stowell, L. 1995. A new disease of poa? PACE Insights 1:3.
29. Stowell, L. J., and Gelernter, W. D. 2001. Diagnosis of Turfgrass Diseases. Ann. Rev. Phytopathol. 39:135-55.
30. Stowell, L. J., and Gelernter, W. 2003. Prograss in understanding rapid blight of cool season turf. PACE Insights 9:3.

