
Fig. 1. Erwin F. Smith's Laboratory in 1905.
Reprinted from Campbell et al. |
What did we learn before 1900?
We learned about blight and mildew in grain in about 520 BC in Israel (Amos 4:9 and Haggai 2:17) but we do not know the specific disease. From then to about the 1700s, however, we learned very little. Further discoveries had to await the development of printing to enable investigators to communicate by means of a written record (which depended on Gutenberg's development of movable type in 1454); the development of a compound microscope to observe microorganisms (by Leeuwenhoek in the 1670s); and still later, a means for culturing microorganisms for study. Needham discovered plant-parasitic nematodes in wheat grains in 1743, Tillet learned that bunt is a disease of wheat in 1755, and Prevost showed that bunt is a fungus in 1807, and gave the first experimental evidence that a disease was caused by a microorganism. Pasteur, the founder of microbiology, formulated the germ theory of disease in 1858 and demolished the theory of spontaneous generation.

Fig.2. Anton deBary.
Reprinted from Campbell
et al. | DeBary, the father of mycology, worked out the life cycle of
Phytophthora infestans in 1853 and showed it to cause late blight of potato, but more importantly established conclusively that fungi are causes not results of disease. Kühn in 1858 established plant disease etiology on a sound scientific basis. Koch developed methods for growing bacteria on liquid and solid media and developed pure culture techniques in 1881 and formulated his famous postulates in 1884. Millardet in 1885 used fungicides (Bordeaux mixture) for disease control in grape, and laid the foundation for plant protection by using chemicals. Burrill discovered bacteria as the cause of fire blight in apple and pear in 1878, and Beijerinck working with tobacco mosaic showed it to be caused by a virus in 1898. So the stage was set for the 1900s. |
What have we learned since 1900?
The milestones of the past century are many and only highlights are described. These highlights have been divided into two categories of ten each: Parasites and Pathogens, and Concepts and Principles.
The top ten in the category of parasites and pathogens, in chronological order, are:
1.
Agrobacterium tumefaciens.
The bacterial nature of crown gall was determined by Smith et al. in 1911 and this finding led eventually, by others, to a tumor-inducing principle, or a tumorigenic substance, a metabolic product of the bacterium. The molecular basis of tumorigenesis was established and the substance was thought to rest in the plasmid DNA molecules of the bacterium. |

Fig. 3. Erwin F. Smith.
Reprinted from Campbell et al. |
2. Tobacco mosaic virus. In 1935, a crystalline substance with properties of tobacco mosaic virus (TMV) was isolated from the juice of TMV-infected tobacco plants by Stanley in 1935, and revealed by use of an electron microscope. The crystals were 100 times more active than a suspension made from TMV-infected tobacco leaves. This was the first report of crystallization of a virus and was followed by extensive research on viruses as causes of plant disease. Classic experiments in virology and etiological biology followed, especially noteworthy was research by Fraenkel-Conrat and Williams in 1955 who isolated protein and RNA from TMV and assembled them to give intact infectious virus particles.
3.
Fusarium oxysporum. The section Elegans (10 species, 18 varieties, 12 forms) in Wollenweber's classification of Fusarium was reduced to a single species,
F. oxysporum, by Snyder and Hansen in 1940, using morphological criteria. He and coworkers went on to further combine species to develop a nine species system of
Fusarium. This work stimulated considerable work with
Fusarium species as pathogens of plants, as well as in work on mycotoxicology in the veterinary and medical sciences.
Fusarium species are parasites or pathogens on nearly all plants, including trees, in all parts of the world, and produce mycotoxins making some plant products hazardous to animals and humans.
4. Mycorrhizae. Although mycorrhizal associations were first defined and named by Frank in 1885, the work by Harley, in 1948 and later, stimulated substantial investigations of rhizosphere fungi as part of the ecology and pathology of root and soil fungi. Later research followed in eliciting physiological defense mechanisms in root infection. The most widespread root infections in plants were probably those caused by vesicular-arbuscular mycorrhizae.

Fig. 4. Ozone (O3) injury symptoms on tobacco.
Reprinted from Schumann. |
5. Air pollutants.Air pollutants were recognized as causal agents of plant disease and gained attention with the report by Middleton and others in 1950, who first reported phytochemical (oxidant) air pollutant effects on plants, especially the effects of smog. The first systematic attempt to estimate the effects of air pollutants on plant productivity was by Benedict et al. in 1971. |
| 6. Phytoplasma and Spiroplasma. Phytoplasmalike organisms were found in preparations from witches' broom of potato and from aster yellows of petunia by Doi et al., in 1967. They observed pleomorphic bodies susceptible to tetracyclines leading them to a phytoplasma hypothesis in the cause of disease. Investigations into other prokaryotes such as the spiroplasmas (discovered by Davis in 1979) and rickettsia-like organisms (Xylella fastidiosa) were stimulated by research into phytoplasma and phytoplasmalike organisms. Phytoplasma lack cell walls. Fundamental to these discoveries was the fact that plant yellows were
not caused by viruses but by phytoplasma of the class Mollicutes. |
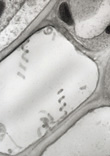
Fig. 5. Spiroplasmas in the phloem of an infected corn plant.
Reprinted from Schumann. |
7. Potato spindle tuber viroid. Small pathogenic RNAs represented a unique class of infectious, low molecular weight molecules and were found to cause potato spindle tuber. These molecules were designated a "viroid" by Diener in 1971, and are almost invisible even when examined with an electron microscope. They have not been found in animals. At least ten diseases are known to be caused by viroids.
8.
Pseudomonas syringae.
With the reduction of all plant pathogenic bacteria to a single species in Bergey's Manual of Systematic Bacteria, a pathovar concept was developed by Young et al. in 1978 and outlined by Dye et al. in 1980, and was subsequently adopted by the International Society for Plant Pathology to complement the International Code of Nomenclature of Bacteria (bacteriological code). This became essential to the identification of bacterial pathogens of plants.
9.
Xylella fastidiosa. Davis in 1978 was the first to isolate the bacterium that causes Pierce's disease of grape. This bacterium was known earlier on as a rickettsia-like organism but is now referred to as a fastidious, Gram-negative, xylem-limited bacterium by Wells et al. (1987). It is not related to the
rickettsiae. |

Fig. 6. Newton B. Pierce.
Reprinted from
Campbell et al. |
10.
Haemophilus influenzae,
Saccharomyces cerevisiae, and
Caenorhabditis elegans. The entire genomes of bacteria (the first prokaryote) in 1995, and yeast (the first eukaryote) in 1996, and the nematode (the first multicellular eukaryote) in 1998 were determined. While these are recent works and they are not plant pathogens, they promise to open genomics as a discipline, and to provide information on the evolution and nature of parasitism in plants and animals, as well as to provide a wealth of new techniques and methodologies.
The top ten in the category of concepts and principles, in chronological order, are:

Fig. 7. Elvin C. Stakman.
Reprinted from Campbell et al. | |
1. The concept of physiologic specialization. Biffen in 1905 was the first to breed crops for disease resistance in cereal rusts using Mendelian principles. But variability in the pathogen was not fully appreciated until the work of Stakman and Piemeisal in 1917 who reported that stem rust in cereals and grasses comprised six biological forms. These forms were distinguished from each other morphologically and parasitically and were differentiated on selected cereal and grass hosts. This led to research on variation and variability of plant pathogens, and subsequently of all microorganisms, and the breeding of plants for resistance to specific races. Races became essential also in microorganisms important in industry, e.g. in the production of acids and enzymes from organisms, and in medicine in the development of antibiotics. Research on the genetics of fungi, the significance of heterokaryosis, the gene-for-gene hypothesis, the development of tolerance and of durable resistance built on work based on the concept of physiologic specialization.
| 2. Controlled environment and disease. Although diseases are caused by pathogens, the environment is a critical factor. To study factors, controlled environments were developed stimulated by work of Johnson in 1921 and 1928 who showed the effects of temperature by using growth chambers. Jones in 1924 had also emphasized the role of environment in causing disease in plants. This led to many sophisticated types of growth chambers and most noteworthy early on was the development of the Pasedena Phytotron described by Went in 1950, which served as a model structure for plant pathologists to use in disease studies. |

Fig. 8. Lewis R. Jones.
Reprinted from Campbell et al. |
3. Field environment in disease and its measurement.
In 1934, Stevens described how Stewart's disease in corn caused by
Aplanobacter stewartii (now
Pantoea stewartii) does not occur when the sum of the mean temperatures from December through February is below 90 but is destructive when the sum exceeds 100, based on 35 years of data. This fostered an appreciation of environmental factors, especially temperature, in the epidemiology of plant disease. Chester in 1950 was the first to look at disease measurement on a quantitative basis in determining losses from disease. Both of these investigations encouraged the development of many models of host-pathogen combinations leading eventually to the mathematical modeling of crop disease as a discipline within plant pathology. A noteworthy compilation of principles including the concept of "r" was developed by Vanderplank in 1963.
4. Phytotoxins. Host-specific toxins were discovered first in two diseases: black spot of Japanese pear by Tanaka in 1933 and in Victoria blight of oat by Meehan and Murphy in 1947. These two diseases became models for further study of phytotoxins. Tanaka reported a heat-labile, host-selective factor produced by
Alternaria kikuchiana that when sprayed on plants produced lesions typical of black spot. A metabolic by-product of
Helminthosporium victoriae that proved toxic to a susceptible oat cultivar was reported by Meehan and Murphy. This heat-stable toxin was found in hyphae and in the growth substrate; symptoms in oat were caused by the toxin. For decades thereafter research continued on the nature and role of phytotoxins in pathogenesis. Most toxins produced by bacteria are not host specific.
5. Phytoalexins.This theory was developed by Müller and coworkers in 1939 and 1940 working with
Phytophthora infestans on potato. The principle is that a substance is present in host tissue that inhibits development of a parasite in hypersensitive tissue and is formed or activated only when the host cells
come in contact with the parasite. Phytoalexins began to serve as an umbrella under which chemical compounds contributing to disease can be classified whether they occur in response to injury, physiological stimuli, infectious agents, or are products of such stimuli or agents. Research led to studies of hypersensitivity and its role in pathology. Although research on phytoalexins has been intensive and stimulated investigations on the nature of resistance to disease, its acceptance as a phenomenon has declined in recent years. Work on disease recognition phenomena, however, emerged to deal with specificity, and the possible role of lectins (Bahlool and Schmidt, 1974).
|
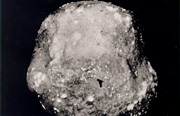
Fig. 9. Potato tubers infected by
Phytophthora infestans.
Reprinted from Schumann.
|
6. Gene-for-gene hypothesis. The pathogenic range in a given race
of Melampsora lini in flax is determined by pathogenic factors specific for each resistant factor in the host according to Flor in 1942. Avirulence is dominant in crosses between races. This work led to the formulation of a gene-for-gene theory by Flor and its application to other host-pathogen combinations in a search for universal applicability. The gene-for-gene concept has been useful in identifying the roles of hybridization, mutation, heterokaryosis, and somatic hybridization in pathogenic variation in parasitic fungi. Interorganismal genetics as described by Loegering in 1978 grew out of Flor's work. Current research delves into molecular characterization in gene-for-gene systems.
7. Organic fungicides. The organic fungicide era was ushered in during 1934 with the introduction of the dithiocarbamates, especially thiram, by Tisdale and Williams of E.I. duPont de Nemours & Company. Iron, lead, and zinc dimethyl derivatives of the carbamates promised the most as fungicides and injured plants the least as reported by Goldsworthy and associates in 1943. Then came the systemic fungicides such as the oxathiins in 1966, and the benzamidazoles in 1966-1968, which raised a new problem of induced resistance.
8. Fungistasis. A fungistatic factor appears to be widespread in soil according to Dobbs and Hinson in 1953. This factor is masked by glucose or by nutrients liberated from microorganisms in soil. This research was further refined by Lingappa and Lockwood in 1963 and later by other plant pathologists and soil microbiologists, who sought to understand fungistasis in relation to soilborne disease. Studies on suppressive soils in relation to root disease incidence grew out of this work on fungistasis and pertained to work on the biocontrol of root disease. Understanding of the ecology of root-infecting organisms was enhanced by research on fungistasis.
9. Soilborne disease and biological control.
Principles of root infection by fungi were developed by Garrett in 1956 in the first of two volumes. His bringing together the findings and conclusions of earlier workers led to his publication on the biology of root-infecting fungi, a monumental work that influenced and stimulated further research on root pathology and the ecology of soil microorganisms and provided a terminology and ecological principles useful in the understanding of biological control of plant pathogens. Hiltner in 1905 had already defined the rhizosphere. Weindling in 1932 was the first to use a known antibiotic-producing antagonist in plant disease control, in reporting parasitism and antagonism of
Trichoderma viride to Rhizoctonia solani. The first commercially successful application of biological control by pitting an organism against a plant pathogen was done by New and Kerr in 1972 in which they dipped seedlings of peach into cell suspensions of
Agrobacterium radiobacter to control crown gall. These investigations on root pathology, soil microbial ecology, and biocontrol applied principles defined by Garrett in his publications.

Fig. 10. Crown Gall (Agrobacterium tumefaciens).
Reprinted from Schumann. | |
10. Gene location in tumor-inducing plasmids.A major change in research on host-parasite interaction took place about 1975 in the physiology and biochemistry of disease when molecular biology and molecular genetics were applied in host-parasite interactions. To determine a possible role of a given molecule in pathogenesis, a gene is cloned and then bits disrupt activity by transformation. The greatest effect of
molecular genetics on plant pathology has been through
Agrobacterium tumefaciens. The first major breakthrough was reported in 1975 when it was established that genes controlling crown gall induction were located in a plasmid, the tumor-inducing plasmid (pTi), by Van Larebke et al. (1975) and Watson et al. (1975). Transposons are discrete sequences of DNA incapable of replication and can insert into replications such as plasmids and chromosomes. Transformation strategies depend on stable introduction of foreign DNA stably into the genome of a cell followed by regeneration of that cell into a whole plant. Virulence by fungi is influenced by dsRNA genetic elements as shown in transmissible hypovirulence in
Cryphonectria parasitica (Nuss and Koltin, 1990).
What have we learned about scientists in the past century?
In the past century considerable changes occurred in the relationships of research workers to universities and institutions. After the appearance of Sputnik, in 1957, there was a major change in funding for research in the United States and in some other nations. Instead of depending on financial support from government, experiment stations, or institutions, investigators had to become entrepreneurs and prepare grant proposals to seek their own funds for doing research. Sometimes, that focused their efforts on what was likely to get funded and precluded research in which the risk of success was unpredictable. In addition, the science of plant pathology, like many other scientific disciplines, became more fragmented as subdisciplines emerged. This fragmentation of the discipline led also to fragmentation of societies and journals, and even to the publication of papers. Whereas, earlier publications represented considerable research, later papers approached a one-experiment, one-paper philosophy, with a consequent proliferation of publications. Then, fax machines and computers became more actively used in research programs, for communication as well as for data processing. Thus specialists at one institution were more likely to communicate with their counterparts throughout the world, than with their colleagues down the hall. These factors contributed to a loss of collegiality and communication among faculty at a given institution, less loyalty to a department or institution, and some erosion of the department as a major force in maintaining a specific discipline. Another major change was the increase in the number of women among scientists. Women brought new talent and new ways of doing things to enrich research programs but this did not alter the factors listed above. Although some departments suffered some as a force in the discipline, other departments maintained their traditional strengths by recruiting specific faculty instead of using the search committee approach. With the advent of molecular biology, there has come to be a blurring of the fields of science and contributions to plant pathology are not always made by plant pathologists.
References
1. Benedict, H. M., Miller, C. J., Olson, R. E. 1971. Economic impact of air pollutants on plants in the United States. Menlo Park, Calif.: Stanford Res. Inst. 77 p.
2. Biffen, R.H. 1905. Mendel's laws of inheritance and wheat breeding. J. Agric. Sci. Camb. 1: 4-48.
3. Bohlool, B. B., and Schmidt, E. L. 1974. Lectins: a possible basis for specificity in Rhizobium-legume nodule symbiosis. Science 185: 269-271.
4. Burrill, T.J. 1880. Pear and apple tree blight. Trans. Ill. State Hortic. Soc. 14: (new Series): 157-167.
5. C. elegans Sequencing Consortium. 1998. Genome sequence of the nematode
Caenorhabditis elegans. A platform for investigating biology. Science 282: 2012-2018.
6. Chester, K. S. 1950. Plant disease losses: their appraisal and interpretation. Plant Dis. Rep. Suppl. 193: 189-362.
7. Davis, M. J., Purcell, A. H., and Thomson, S. V. 1978. Pierce's disease of grapevines: isolation of the causal bacterium. Science 199: 75-77.
8. Davis, R. E. 1979. Spiroplasma, newly recognized arthropod-borne pathogens. In Leafhopper Vectors and Plant Disease Agents, ed. K. F. Harris, K. Maramorosch. pp 451-488. NT Acad. Press.
9. Diener, T.O. 1971. Potato spindle tuber 'virus' IV. A replicating, low molecular weight RNA. Virology 45: 411-428.
10. Dobbs, C.G., and Hinson, W.H. 1953. A widespread fungistasis in soils. Nature 172: 197-198.
11. Doi, Y., Teranaka, M., Yora, K., and Asuyama, H. 1967. Mycoplasma- or PLT group-like microorganisms found in the phloem elements of plants infected with mulberry dwarf, potato witches' broom, aster yellows, or Paulownia witches' broom. Ann. Phytopathol. Soc. Jpn. 33: 259-266.
12. Dye, D. W., Bradbury, J. F., Goto, M., Hayward, A. C., Lelliot, R. A., and Schroth, M. N. 1980. International standards for naming pathovars of phytopathogenic bacteria and a list of pathovar names and pathogenic strains. Rev. Plant Pathol. 59: 153-168.
13. Fleischmann, R. D., Adams, M. D., White, O., Clayton, R.A., Kirkness, E.F., Kerlavage, A. R., Bult, C. J., Tomb, J. F., Dougherty, B. A., and Merrick, J. M., et al. 1995. Whole-genome random sequencing and assembly of
Haemophilus influenzae Rd. Science 269: 496-512.
14. Flor, H. H. 1942. Inheritance of pathogenicity in
Melampsora lini. Phytopathology 32: 653-669.
15. Fraenkel-Conrat, H., and Williams, R.C. 1955. Reconstitution of active tobacco mosaic virus from its inactive protein and nucleic acid components. Proc. Nat. Acad. Sci. USA 41: 690-698.
16. Frank, A.B. 1885. Öber die auf Wurzelsymbiose beruhende Ernahrung gewissor Baume durch unterirdische Pilze. Ber. Deut. Botan. Ges. 3:128-145.
17. Garrett, S.D. 1956. Biology of root-infecting fungi. Cambr. Univ. Press. 293 p.
18. Goffeau, A., Barrell, B. G., Bussy, H., Davis, R. W., Dujon, B., Feldman, H., Galibert, F., Hoheisel, J.D., Jacq, C., Johnston, M. et al. 1996. Life with 6000 genes. Science 274: 546-567.
19. Goldsworthy, M.C., Green, E.L., and Smith, M.A. 1943. Fungicidal and phytocidal properties of some metal dialkyl dithiocarbamates. J. Agric. Res. 66: 277-291.
20. Harley, J.L. 1948. Mycorrhiza and soil ecology. Biol. Rev. 23: 127-158.
21. Johnson, J. 1921. The relation of air temperatures to certain plant diseases. Phytopathology 11: 446-458.
22. Johnson, J. 1928. Constant temperature and humidity chambers. Phytopathology 18: 227-238.
23. Jones, L. R. 1924. The relation of environment to disease in plants. Am. J. Bot. 11: 601-609.
24. Lingappa, B.T., and Lockwood, J.L. 1961. The nature of the widespread soil fungistasis. J. Gen. Microbiol. 26: 473-485.
25. Loegering, W.Q. 1978. Current concepts in interorganismal genetics. Ann. Rev. Phytopathol. 16: 309-320.
26. Meehan, F., and Murphy, H.C. 1947. Differential phytoxicity of metabolic by-products of
Helminthosporium victoriae. Science 106: 270-271.
27. Middleton, J. T., Kendrick, J. B. Jr., Schwalm, H. W. 1950. Injury to herbaceous plants by smog or air pollution. Plant Dis. Rep. 34: 245-252.
28. Müller, K.O., and Berger, H. 1940. Experimentelle Untersuchungen öber die Phytophthora-Resistenz der Kartoffel. Arb. Biol. Reichsanstalt Land-u. Forstwirtsch., Berlin 23: 189-231.
29. New, P.B., and Kerr, A. 1972. Biological control of crown gall: field measurements and glasshouse experiments. J. Appl. Bacteriol. 35: 297-287.
30. Nuss, D.L., and Keltin, Y. 1990. Significance of dsRNA genetic elements in plant pathogenic fungi. Ann. Rev. Phytopathol. 28: 37-58.
31. Smith, E. F., Brown, N. A., Townsend, C. O. 1911. Crown-gall of plants: Its cause and remedy. USDA Bur. Plant Ind. Bull. 213: 1-200.
32. Snyder, W.C., and Hansen, H.N. 1940. The species concept in
Fusarium. Am. J. Bot. 27:64-67.
33. Stakman, E.C., and Piemeisal, F.J. 1917. Biologic forms of
Puccini graminis on cereals and grasses. J. Agric. Res. 10: 429-495.
34. Stanley, W.M. 1935. Isolation of a crystalline protein possessing the properties of tobacco-mosaic virus. Science 81: 644-645.
35. Stevens, N.E. 1934. Stewart's disease in relation to winter temperatures. Plant Dis. Rep. 18: 141-149.
36. Tanaka, S. 1933. Studies on the black spot disease of the Japanese pears (Pyrus serotina). Mem. College Agr. Kyoto Imp. Univ. 28: 1-31.
37. Vanderplank, J. 1963. Plant Diseases: Epidemics and Control. Acad. Press, NY. 349 p.
38. Van Larebke, N., Genetello. C., Schell, J., Schilperoot, RA., Hermans, A.K. et al. 1975. Acquisition of tumour-inducing ability by the non-oncogenic agrobacteria as a result of plasmid transfer. Nature 255: 742-743.
39. Watson, B., Currier, T. C., Gordon M.P., Chilton M.-D., Nester, E.W. 1975. Plasmid required for virulence of
Agrobacterium tumefaciens. J. Bacteriol. 123: 255-264.
40. Weindling, R. 1934. Studies on the lethal principle effective in the parasitic action of
Trichoderma lignorum on
Rhizoctonia solani and other soil fungi. Phytopathology 24: 1153-1179.
41. Wells, J.M., Raju, B.C., Hung, H.Y., Weisburg, W. G., Mandelco-Paul, L., Brenner, D. J. 1987.
Xylella fastidiosa gen. nov. sp. nov.: Gram-negative, xylem-limited, fastidious plant bacteria related to
Xanthomonas spp. Int. J. Syst. Bacteriol. 37: 136-143.
42. Went, F. 1950. The Earhart Plant Research Laboratory. Chron. Bot. 12: 91-108.
43.Young, J. M., Dye, D.W., Bradbur, J. F., Panagopoulos, C. G., and Robbs, C.F. 1978. A proposed nomenclature and classification for plant pathogenic bacteria. NZ J. Agric. Res. 21: 153-177.
Figures
Figures 1,3,6,7, and 8 reprinted from
The Formal Years of Plant Pathology in the United States by Campbell, Peterson, and Griffith. Copyright 1999 The American Phytopathological Association.
Figures 2,4,5,9, and 10 reprinted from
Plant Diseases: Their Biological and Social Impact by Gail Schumann.
Copyright 1991 The American Phytopathological Association.

