:
Anthracnose of turfgrass
Colletotrichum cereale (formerly Colletotrichum graminicola) At least thirteen warm and cool-season turfgrass species are hosts. However, anthracnose is predominantly a disease of annual bluegrass (Poa annua) and bentgrass (Agrostis spp.) grown as managed turf.
Authors
Derek M. Settle, Alfredo D. Martinez-Espinoza, and Lee L. Burpee
University of Georgia, Griffin Campus, Griffin, GA

Reddish brown foliar lesions on creeping bentgrass caused by Colletotrichum cereale.
Symptoms and Signs
Symptoms
Anthracnose of turfgrass is a foliar disease or a basal rot of the lower stem. Foliar lesions on creeping bentgrass appear reddish-tan (Figure 1). In some cases discrete leaf spot symptoms may not develop, and instead, older leaf blades are completely water-soaked (Figure 2). The foliar phase of anthracnose is also common on annual bluegrass where symptoms include elongate, chlorotic (yellow) leaf spots with small, black, spore-bearing structures (acervuli) in the center (Figure 3). The leaves turn brown and may be peppered with acervuli (Figure 4).
Figure 1 |

Figure 2 |

Figure 3 |

Figure 4 |
Basal rot occurs when leaf sheaths at the base of tillers are colonized by the fungus. Infected tissue appears dark brown to black, and leaf blades appear orange to yellow and are devoid of lesions (Figure 5). Creeping bentgrass stolons may also become infected (Figure 6).

Figure 5 |

Figure 6 |
An anthracnose outbreak in a golf putting green, tee, or fairway can have a patchy (Figures 7, 8) or diffuse (Figure 9) appearance. Foci of diseased plants can range from small irregular patches that measure 1 to 10 cm (>0.5 to 4 in.) to large areas that measure >1 m (> 3 ft). Severe outbreaks often result when the pathogen selectively kills invading annual bluegrass in creeping bentgrass putting greens (Figures 9, 10). Anthracnose can also develop on creeping bentgrass putting greens that have little or no annual bluegrass infestation (Figure 11). Damage to creeping bentgrass can progress to cause significant plant thinning within individual patches. The reason for patch formation is uncertain, but some patches may represent susceptible biotypes of bentgrass that segregate from more resistant biotypes within a putting green (Figure 12).

Figure 7 |

Figure 8 |

Figure 9 |

Figure 10 |

Figure 11 |

Figure 12 |
Signs
The most prominent sign of anthracnose is the presence of tufts of black setae (sterile, hair-like fungal structures) on plant tissue. Setae can be observed with a 10X magnification lens and are often abundant on dead or dying leaves, leaf sheaths, or stolons (Figure 13). Setae denote the presence of acervuli (Figure 14), fruiting bodies which produce asexual spores (conidia) contained in a mucilaginous matrix (Figure 15). The mucilaginous material prevents desiccation damage and premature germination of conidia. Conidia are hyaline (clear), crescent-shaped, and single celled (Figure 16). Conidia may be confused with those of Fusarium spp. and Microdochium nivale (cause of pink snow mold), but these species usually form conidia that have two or more cells.

Figure 13 |

Figure 14 |

Figure 15 |
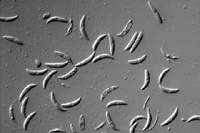
Figure 16 |
Pathogen Biology
Colletotrichum cereale overwinters as mycelium (Figure 17) or conidia associated with previously infected plant tissue. The fungus may also survive as darkly pigmented aggregates of hyphal cells (stromata) that are formed on stolons and at the base of tillers (Figure 18). Exposure of the stromata to sunlight and moderate temperatures of 15 to 25ºC (59 to 77ºF) can induce formation of conidia. The conidia may serve as initial inoculum for basal rot anthracnose in the spring or early summer. However, further research is required to determine whether basal rot anthracnose and foliar anthracnose are two distinct diseases caused by different strains of C. cereale or simply phases of a single disease. Conidia are spread to surrounding leaf blades by water, mowing, and other cultural practices that cause physical disruption of the turfgrass sward.

Figure 17 |
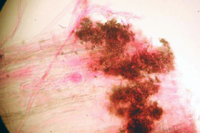
Figure 18 |
In early to mid-summer during warm, wet periods, conidia may also germinate on plant surfaces to form germ tubes that develop into lobed, melanized appressoria (Figure 19). Appressoria serve to anchor the fungal filaments to the plant surface. A penetration peg emerges from a pore in the appressorium and penetrates plant epidermal cells directly. Penetration of the epidermis is closely followed by production of a single, irregularly-shaped infection hypha. One or more branches from the infection hypha then invade surrounding plant mesophyll cells. From this point, C. cereale behaves as a necrotroph, killing plant tissue with enzymes, and possibly toxins, in advance of mycelium growth. The sexual stage (teleomorph) of C. cereale is the ascomycete Glomerella graminicola. This stage has been observed in laboratory cultures but not on infected turfgrasses.
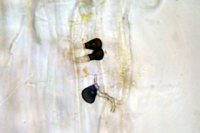
Figure 19 |
Disease Cycle and Epidemiology
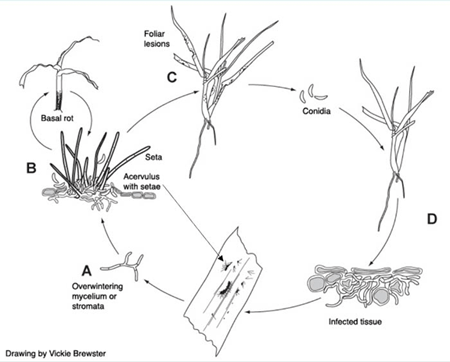
A) Colletotrichum cereale overwinters as mycelium or stromata associated with stolons and stems in thatch. B) In spring, infection begins on leaf sheaths and stolons. This early infection can result in symptoms of anthracnose basal rot. C) Multiple acervuli on infected turfgrass foliage produce conidia that are splashed or displaced from leaves to infect healthy turfgrass plants. Anthracnose foliar blight causes thinning of turfgrass during early to mid-summer. D) The fungus penetrates epidermal cells and colonizes mesophyll cells. Yellow foliar lesions and black acervuli develop.
Epidemiology
Anthracnose develops over a range of temperatures. Symptoms of basal rot develop at cooler temperatures (15 to 25°C, or 60 to 78°F), and foliar symptoms occur at temperatures above 26°C (79°F). Anthracnose foliar blight outbreaks have been predicted by using a combination of leaf wetness duration and temperature. Continuous leaf wetness of 12 hours or more and temperatures of 14 to 28°C (57 to 82°F) are necessary for infection, which intensifies with increasing temperature and leaf wetness duration. The latent period (the time between infection and production of new spores) is thought to be 10 to 12 days.
Juvenile plants display good resistance to foliar blight caused by C. cereale. As plants mature, initial symptoms develop on older, senescing leaves. In annual bluegrass, foliar symptoms of anthracnose increase in severity following anthesis (opening of flowers). Plants subjected to drought and/or fertility stress are believed to be more susceptible to infection.
Host specificity. Isolates of C. cereale show a degree of host specificity to either annual bluegrass or creeping bentgrass, although some isolates can infect both turfgrass hosts. DNA analysis indicates that isolates from annual bluegrass, creeping bentgrass, and corn are genetically different. Furthermore, genetic similarities among C. cereale isolates are influenced more by plant host than by geographic origin. For example, isolates of C. cereale from creeping bentgrass and annual bluegrass may differ in the length of both appressoria and spores. Results of one study indicated that mean appressorial length was approximately 0.8 µm longer for creeping bentgrass isolates than for annual bluegrass isolates. In contrast, mean spore length was 1.5 µm shorter for creeping bentgrass isolates than for annual bluegrass isolates. In addition, when cultured on agar media, the color of C. cereale colonies was influenced by turfgrass host. Black-gray cultures were isolated from creeping bentgrass, and cultures with pink coloration were isolated from annual bluegrass.
Disease Management
Cultural management
Susceptibility to Colletotrichum cereale is enhanced in turfgrass that is subjected to stresses such as high temperature, root zone compaction, thatch accumulation, low fertility, drought, low mowing height, and insect infestation. Cultural practices that reduce these and other stresses often help to limit anthracnose incidence and severity. These practices include core aerification, dethatching, topdressing, use of light weight equipment including mowers, increased nitrogen fertilization, and sufficient irrigation to prevent drought stress. To be effective, however, such management practices should be initiated before anthracnose symptoms develop. Some cultural practices such as core aerification, dethatching, and topdressing may create wounds that serve as infection sites for C. cereale. Therefore, a fungicide treatment may be required prior to use of these procedures. Since spores of C. cereale require more than 12 hours of continuous leaf wetness to germinate and infect, attempts should be made to avoid long periods of leaf wetness. Leaf surfaces should be allowed to dry completely before sunset. Trees and shrubs should be pruned to enhance air circulation, and fans can be strategically placed around turf areas where air circulation is poor.
Chemical management
Several contact (e.g., chlorothalonil), penetrant (e.g., trifloxystrobin), and systemic (e.g., propiconazole) fungicides are registered for control of anthracnose. Each of these fungicides can provide preventive (pre-infection) control, whereas only penetrant and systemic fungicides have curative (post-infection) activity. In some states of the US, populations of C. cereale have developed resistance to benzimidazole (e.g., thiophanate methyl) and QoI (quinone outside inhibitor) fungicides (e.g., azoxystrobin, trifloxystrobin, and pyraclostrobin).
Biological control
Surface hydrophobicity has been found to induce spore germination of C. cereale on corn. In the future, selection of annual bluegrass and creeping bentgrass with a non-waxy cuticle may be a novel strategy that can delay or interfere with germination of C. cereale conidia, as has been suggested for corn.
Significance
Historical Significance
In the United States, the fungus now known as Colletotrichum cereale was first reported in association with bluegrass in 1914. In 1928, a disease of Poa annua with three causal fungal organisms was described. The organisms described are now known as C. cereale, Drechslera poae, and Fusarium sp. The relative importance of the three fungi in causing the disease was not known. Experiments in England during the 1950s suggested that C. cereale was responsible for the basal rot symptoms of annual bluegrass.
In the late 1960s, anthracnose was described as an important foliar disease of annual bluegrass in South Carolina. However, the existence of the disease was challenged, based on the observation that C. cereale infection was associated with hot environmental conditions and senescing turfgrass leaf blades. Prior to 1975, summer mortality of annual bluegrass was considered a physiological disorder, and plant death was attributed to mid-day heat stress. In 1976, anthracnose was proposed to be one of three components of a syndrome called “Helminthosporium anthracnose senescence” (HAS) decline. The defined syndrome reflected the association of C. cereale with leaf spotting fungi (Bipolaris or Drechslera) and senescing older leaves. By the mid 1980s, C. cereale was confirmed to be pathogenic to annual bluegrass in the United States by fulfilling Koch’s postulates.
Prior to the 1970s, anthracnose was reported sporadically on golf putting greens, but the disease has become more prevalent in recent decades. The increase of anthracnose epidemics on golf courses in the United States coincides with the use of cultural practices that induce greater stress on turfgrass plants, such as reducing mowing height, limiting fertilizer applications, and growing turf in compacted soil.
Selected References
Backman, P. A., P. J. Landschoot, and D. R. Huff. 1999. Variation in pathogenicity, morphology, and RAPD marker profiles in Colletotrichum graminicola from turfgrasses. Crop Science 39:1129-1135.
Bergstrom, G. C. and R. L. Nicholson. 1999. The biology of corn anthracnose. Plant Disease 83:596-608.
Bolton, A. T. and W. E. Cordukes. 1981. Resistance to Colletotrichum graminicola in strains of Poa annua and reaction of other turfgrasses. Canadian Journal of Plant Pathology 3:94-96.
Browning, M., L. V. Rowley, P. Zeng, J. M. Chandlee, and N. Jackson. 1999. Morphological, pathogenic, and genetic comparisons of Colletotrichum graminicola isolates from Poaceae. Plant Disease 83:286-292.
Burpee, L. 2005. Sensitivity of Colletotrichum graminicola to phosphonate fungicides. International Turfgrass Society Research Journal 10:163-169.
Chaky, J., K. Anderson, M. Moss, and L. Vaillancourt. 2001. Surface hydrophobicity and surface rigidity induce spore germination in Colletotrichum graminicola. Phytopathology 91:558-564.
Couch, H. B. 1995. Diseases of Turfgrasses, 3rd edition. Krieger Publishing Company. Malabar FL.
Crouch, J. A., B. B. Clarke, and B. I. Hillman. 2006. Unraveling evolutionary relationships among the divergent lineage of Colletotrichum causing anthracnose disease in turfgrass and corn. Phytopathology 96:46-60.
Danneberger, T. K., J. M. Vargas, Jr., P. E. Rieke, and J. R. Street. 1983. Anthracnose development on annual bluegrass in response to nitrogen carriers and fungicide application. Agronomy Journal 75:35-38.
Danneberger, T. K., J. M. Vargas, Jr., and A. L. Jones. 1984. A model for weather-based forecasting of anthracnose on annual bluegrass. Phytopathology 74:448-451.
Horvath, B. J. and J. M. Vargas Jr. 2004. Genetic variation among Colletotrichum graminicola isolates from four hosts using isozyme analysis. Plant Disease 88:402-406.
Jackson, N. and V. J. Herting. 1985. Colletotrichum graminicola as an incitant of anthracnose/basal stem rotting of cool season turfgrasses. Proceedings Fifth International Turf Research Conference 5:647-656.
Khan, A. and T. Hsiang. 2003. The infection process of Colletotrichum graminicola and relative aggressiveness on four turfgrass species. Canadian Journal of Microbiology 49:433-442.
Mims, C. W. and L. J. Vaillancourt. 2002. Ultrastructural characterization of infection and colonization of maize leaves by Colletotrichum graminicola, and by a C. graminicola pathogenicity mutant. Phytopathology 92:803-812.
Smiley, R. W., P. H. Dernoeden, and B. B. Clarke. 2005. Compendium of Turfgrass Diseases, 3rd ed. American Phytopathological Society, St. Paul, MN.
Smith, J. D., N. Jackson, and A. R. Woolhouse. 1989. Fungal Diseases of Amenity Turfgrasses. E. and F. N. Spon Ltd., New York, NY.
Sutton, B. C. 1968. The appressoria of Colletotrichum graminicola and C. falcatum. Canadian Journal of Botany 46:873-876.
Vargas, J. M., Jr. and R. Detweiler. 1985. Anthracnose of Poa annua: the pathogenicity of Colletotrichum graminicola. International Turfgrass Society Research Journal 5:637-640.
Vargas, J. M., Jr., T. K. Danneberger, and A. L. Jones. 1993. Effects of temperature, leaf wetness duration, and inoculum concentration on infection of annual bluegrass by Colletotrichum graminicola. International Turfgrass Society Research Journal 6:324-328.
Vargas, J. M., Jr. 1994. Management of Turfgrass Diseases, 2nd edition. Lewis publishers, Boca Raton, FL.

