Bacterial leaf scorch (BLS) of shade trees
Xylella fastidiosa
Bacterial leaf scorch (BLS) affects many different shade tree species (Table 1) such as American elm, red maple, sweet gum, sycamore and London plane, and a number of species of oak (Figure 1). The disease has been identified in the urban forest (landscapes, street plantings, and small woodlots) throughout the eastern United States and as far west as Texas.

Shade tree hosts affected by BLS
Table 1. Shade tree hosts affected by BLS.
|
Scientific name |
Common name |
|
Acer sp. | |
| A. rubrum | Red maple |
| A. negundo | Boxelder |
| A. saccharum | Sugar maple |
|
Cornus florida | Flowering dogwood |
|
Celtis occidentalis | Hackberry |
|
Liquidambar stryraciflua | Sweet gum |
|
Morus alba | White mulberry |
|
Platanus sp. | |
| P. occidentalis | American sycamore |
| P. x acerifolia | London plane |
|
Quercus sp. | |
| Q. velutina | Black oak |
| Q. incana | Bluejack oak |
| Q. macrocarpa | Bur oak |
| Q. prinus | Chestnut oak |
| Q. laurifolia | Laurel oak |
| Q. virginiana | Live oak |
| Q. rubra | Northern red oak |
| Q. palustris | Pin oak |
| Q. stellata | Post oak |
| Q. coccinea | Scarlet oak |
| Q. imbricaria | Shingle oak |
| Q. shumardii | Shumard oak |
| Q. falcata | Southern red oak |
| Q. bicolor | Swamp white oak |
| Q. laevis | Turkey oak |
| Q. nigra | Water oak |
| Q. alba | White oak |
| Q. phellos | Willow oak |
|
Ulmus americana | American elm |
Table 2. Some of the economically important diseases caused by
Xylella fastidiosa characterized by the primary symptom expressed.
|
Leaf scorch | Almond leaf scorch (Prunus amygdalus) |
| Bacterial leaf scorch of shade trees |
| Coffee leaf scorch (Coffea arabica) |
| Oleander leaf scorch (Nerium oleander) |
| Pear leaf scorch (Pyrus pyrifolia) |
| Pecan leaf scorch (Carya illinoinensis) |
| Pierce’s disease of grapevine (Vitis spp.) |
| Plum leaf scald (Prunus domestica,
P. salicina) |
|
Stunt | Alfalfa dwarf (Medicago sativa) |
| Citrus variegated chlorosis (Citrus spp.) |
| Phony peach disease (Prunus persica) |
| Periwinkle wilt (Catharanthus roseus) |
Table 3. Some alternative hosts of
Xylella fastidiosa.1
|
Scientific name |
Common name |
|
Aesculus x hybrid | Buckeye |
|
Ampelopsis arborea | Peppervine |
|
Ampelopsis brevipedunculata | Porcelain berry |
|
Artemisia spp. | Mugwort |
|
Baccharis halimifolia | Eastern baccharis |
|
Callicarpa americana | American beautyberry |
|
Celastrus orbiculata | Oriental bittersweet |
|
Cynodon dactylon | Bermuda grass |
|
Fagus crenata | Japanese beech bonsai |
|
Fragaria californica | Wild strawberry |
|
Hedera helix | English ivy |
|
Montia linearis | Miner’s lettuce |
|
Parthenocissus quinquefolia | Virginia creeper |
|
Parthenocissus tricuspidata | Boston ivy |
|
Paspalum dilatatum | Dallis grass |
|
Rhus sp. | Sumac |
|
Rubus procerus | Blackberry |
|
Sambucus canadensis | American elder |
|
Solidago fistulosa | Goldenrod |
|
Sorghum halapense | Johnson grass |
|
Trifolium repens var.
latum | Landino clover |
|
Vitis sp. | Wild grape |
For a more complete list of alternative hosts, refer to the
Xylella fastidiosa web site:
http://www.cnr.berkeley.edu/xylella/index.html.
Modified from the Nov 2005 APSnet Feature Article,
http://publish.apsnet.org/publications/apsnetfeatures/Pages/BacterialLeafScorch.aspx
Symptoms and Signs
Leaf scorching in shade trees
Leaf scorch in plants can be attributed to biotic agents or abiotic stresses. Abiotic (or environmental) stresses that can cause leaves to scorch include moisture extremes, wind, salt, air pollutants, toxic metals, and nutrient extremes. In most cases, this type of scorching is fairly uniform around leaf edges, affects newer leaves as well as older leaves, will appear on vast expanses of the canopy, and may also develop soon after a known stress (such as drought or an application of salt) occurs (Figure 2).

Figure 2 |
Plant infection by living or biotic agents (such as fungi, bacteria, nematodes, and viruses) can also result in leaf scorching, but this type of scorching is not clearly defined on the plant. Symptoms on leaves are often irregular in shape, and frequently a yellow or red "band" will appear between green and scorched tissues (Figure 3). In addition, symptoms may appear first on leaves of one or more branches, and then, over time, appear on other parts of the tree.

Figure 3 |
Symptoms.Symptoms of BLS vary by shade tree host (Figure 4), but in most cases the disease is identified by a characteristic marginal leaf scorch similar to that described above for biotic agents.

Figure 4a |

Figure 4b |

Figure 4c |

Figure 4d |
Symptoms first appear in late summer to early fall. In trees with determinate growth, such as oak, the scorching appears on leaves of all ages at about the same time. In trees with indeterminate growth, such as sycamore and elm, symptoms progress from older to younger leaves. Affected leaves may curl and drop prematurely, and as the disease progresses over several years, branches die and the tree declines. Elms may be killed outright by the disease; other affected species eventually decline to the point where the dead branches pose a risk and the tree must be removed. The process of tree decline may occur quickly or slowly, perhaps depending on the host and the environment. Epicormic sprouts (shoots that arise from adventitious or latent buds) may be prominent on severely diseased trees, and scale insects, borers, Armillaria root rot, and other biotic diseases may be present as secondary pests.
The symptoms and distribution of several important shade tree hosts affected by BLS are described below.
Elm. Leaves of elm (Ulmus americana) affected by BLS have an irregular pattern of necrosis (tissue death) along the margin that is often accompanied by a chlorotic (yellow) halo (Figure 3). This irregular scorch is quite different from the uniform pattern caused by environmental stress (such as drought). Symptoms progress from older to younger leaves (Figure 5).

Figure 5 |
Elms affected by BLS are also very susceptible to
Dutch elm disease (DED) (which is the usual reason such trees die and are removed). DED differs from BLS, however, in that leaves associated with DED flag and curl (not scorch) earlier in the growing season, and xylem discoloration, which is a diagnostic characteristic for DED, does not occur in trees affected by BLS. BLS of elm has been detected as far north as the Niagara Peninsula and is particularly troublesome in the mid-Atlantic United States. For example, in 2001, 30% of 3000 elm trees planted in the monumental core in Washington, D.C. were affected by the disease.
Sycamore. BLS of sycamore (Platanus occidentalis) is a chronic disease, and it may be years before trees affected by the disease die. Symptoms appear late in the season as a papery, interveinal leaf scorch with a narrow chlorotic halo (Figure 6). As in elm, the disease progresses from older to younger leaves (Figure 7). BLS of sycamore may be confused with sycamore anthracnose, which appears earlier in the growing season and tends to affect tissue along the veins instead of between them.
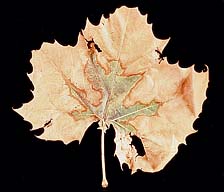
Figure 6 |

Figure 7 |
Oak. As mentioned previously, BLS affects many species of oak. Symptoms of BLS on the leaves of red oak (Quercus rubra) appear as a pronounced, marginal discoloration with a dull red or yellow halo between scorched and green tissues (Figure 1). Unlike elm, however, most, if not all, of the leaves on an affected branch will scorch (Figure 8). This is due to the determinate growth habit (all leaves on a branch share the same age) of oak trees. In the early stages BLS, portions of the tree remain unaffected, while other branches exhibit typical leaf scorch symptoms. As the disease progresses, more branches develop symptoms (Figure 9). Within populations of trees, disease incidence usually appears randomly; trees neighboring severely affected trees are often asymptomatic (Figure 10).

Figure 8 |

Figure 9 |

Figure 10 |
Leaf symptoms in pin oak (Q. palustris) are not as distinct (Figure 11) and can be easily confused with abiotic stresses, but the distribution of the disease within the canopy and between trees is the same as for red oak.

Figure 11 |
BLS of oak may be confused with oak wilt, another vascular disease. Like BLS, initial symptoms of oak wilt appear as scorched leaves. Unlike BLS, however, scorching appears in spring to early summer, and trees defoliate and die within several months after symptoms appear.
BLS of oak is has been reported from New Jersey as far south as Tallahassee, FL, the mid-western states, and in Texas. In some New Jersey municipalities, BLS is known to affect up to 35% of oaks planted as street trees and in landscapes.
Diagnosis
Xylella fastidiosa was not recognized as a pathogen of landscape trees until the early 1980s. Since its symptoms are very similar to those caused by abiotic stresses, it is not surprising that the disease is frequently overlooked or misdiagnosed.
Preliminary diagnosis of BLS is made by interpreting the symptoms described above in late-summer and early fall. Especially useful diagnostic criteria include leaf scorch, premature leaf drop (Figure 12), the random distribution of affected branches around the canopy, thinning of the crown (Figure 13), and the random appearance of the disease within populations of trees.

Figure 12 |

Figure 13 |
Definitive diagnosis is most often made in the laboratory using a selective antibody technique known as enzyme-linked immunosorbent assay (ELISA) on symptomatic leaves to detect the pathogen (Figure 14). Other techniques used to detect
X. fastidiosa include light and electron microscopy, standard and real-time PCR techniques, and immunomagnetic separation followed by PCR (Table 4) Among these methods, ELISA remains the most rapid and cost-effective method to detect the pathogen in symptomatic tissues. The molecular techniques are more sensitive, however, and are helpful for detecting low populations of bacterial cells in infected, asymptomatic hosts or in insect vectors.

Figure 14 |
Table 4. Methods used to detect
Xylella fastidiosa1.
|
Technique |
Sensitivity2
(number of bacterial cells) |
Cost and labor |
| Culture from xylem fluid on selective media | 1000 | Low |
| ELISA3 | 100,000 | Med |
| PCR | 100 | High |
| IC-PCR4 | <100 | High |
1Adapted from R. Jordan. 2002. In: Bacterial Leaf Scorch in Amenity trees: A Wide-Spread Problem of Economic Significance to the Urban Forest. Lashomb, J., A. Iskra, A.B. Gould, and G. Hamilton, eds. Vol. NA-TP-01-03: USFS
2Lowest approximate number of bacterial cells that the assay can detect.
3Enzyme-linked immunosorbent assay
4Immunocapture-PCR
Pathogen Biology
Pathogen morphology
Xylella fastidiosa is a Gram-negative, rod-shaped bacterium that lacks flagella for motility and requires oxygen for respiration. The bacterial cells often possess a rippled (undulating) cell wall and terminal fimbriae (surface structures, shorter than flagella, that help to anchor the cells together in the xylem stream) (Figures 15 and 16).
X. fastidiosa, as the name suggests, has fastidious nutrient requirements and can be difficult to grow in culture. The bacterium grows slowly on selective medium to form small colonies that appear white to yellow (Figure 17). The entire genome of a strain of
X. fastidiosa isolated from citrus has been recently sequenced (the first bacterial plant pathogen for which this has been performed). Based on what is known for other microbes, the
X. fastidiosa genome encodes for proteins involved in cell-cell interaction, degradation of plant cell walls, synthesis of toxins, and ability to cause disease (pathogenicity).

Figure 15 |
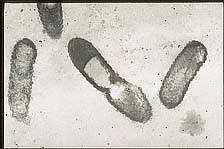
Figure 16 |

Figure 17 |
Host specificity
When the bacterium was first described in 1987, 25 bacterial strains isolated from 10 different hosts were included as a single species. Strains differ, however, in characteristics such as host range, pathogenicity, nutritional requirements, and genetic homology. In 2004, several new subspecies of
X. fastidiosa were proposed based on pathogenicity, phylogenetic characteristics, and DNA relatedness. For example, the bacterium that causes Pierce’s disease of grapevine is
X. fastidiosa subsp.
piercei. Placed in the
X. fastidiosa subsp.
multiplex taxon are strains of
X. fastidiosa that affect elm, sycamore, oak, and maple. Even within the same subspecies, however, differences in host range exist. For example, strains isolated from elm are not pathogenic to sycamore, and
vice versa.
Disease Cycle and Epidemiology
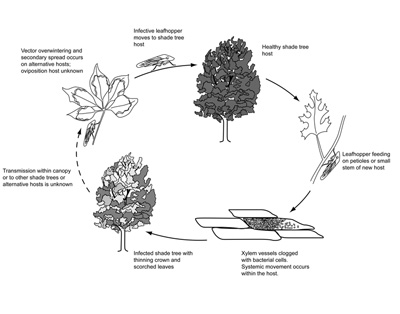
Disease Cycle |
Xylella fastidiosa is spread primarily by leafhopper insects (subfamily Cicadellidae) known as sharpshooters, and to a lesser extent, spittlebugs (family Cercopidae). These insects have piercing-sucking mouthparts and subsist on xylem fluid. Both adult and immature (nymph) stages acquire the bacterium when feeding on succulent tissues of infected hosts. As xylem fluid is drawn into the insect, bacterial cells attach to the cibarial pump and the lining of the esophagus (collectively known as the foregut of the insect). There, the bacterium multiplies and forms a biofilm, where it becomes encased and protected in a bacterial “glycocalyx” (made of polysaccharide and protein), extracting nutrients from xylem fluid as it is pumped through the insect.
Once an insect acquires the bacterium, transmission to a new host can begin within 1 to 2 hours. In the early stages of feeding, bacterial cells become dislodged and are pumped directly into the xylem where systemic movement within the host occurs. An adult can transmit the bacterium for the remainder of its life, whereas nymphs, which shed the foregut during molting, can do so only until the next molt. In grapes with Pierce’s disease, a threshold population of bacteria (104 to 105 colony forming units per gram) is needed before transmission by insect vectors becomes likely. It is not known whether a similar threshold is needed for the pathogens that cause BLS in shade trees.
Xylem-feeding insects, particularly leafhoppers, can be polyphagous (i.e., they feed on many different hosts within a single season). Many of the alternative (non economically important) hosts of
X. fastidiosa mentioned earlier serve as a food source for potential leafhopper vectors, and many leafhoppers overwinter as adults on these alternative plants. Alternative hosts may be the source of a substantial amount of inoculum that is transmitted to economically important crops, such as grape and peach, by vectors that feed on both types of hosts. The insects that vector some economically important diseases (Table 5), such as Pierce’s disease of grapevine and phony peach disease, are known (e.g., species of the leafhoppers
Graphocephala (Figure 18),
Homalodisca, and Oncometopia), and their role in the disease process is well characterized. Insects that vector BLS in shade trees, however, have yet to be identified, nor yet is known the role that alternative hosts play in the disease process for shade tree hosts. Current research on BLS in oak and other shade tree hosts indicates that several known vectors of other diseases caused by
X. fastidiosa are present in shade trees during the growing season (e.g.,
Graphocephala and
Oncometopia species). Their role in the transmission of BLS has yet to be confirmed.

Figure 18 |
Table 5. Some known leafhopper vectors of
Xylella fastidiosa.
|
Host |
Insect vector |
| Citrus (Brazil) |
Acrogonia terminalis
Dilobopterus costalimai
Oncometopia fascialis
Oncometopia nigricans |
| Grape |
Carneocephala fulgida
Draeculacephala minerva
Graphocephala atropunctata
Homalodisca coagulata
Oncometopia nigricans |
| Oleander |
Homalodisca coagulata
Homalodisca lacerta |
| Peach, plum |
Graphocephala versuta
Homalodisca coagulata
Homalodisca insolita
Oncometopia orbona |
In some hosts,
X. fastidiosa is known to pass through root grafts (e.g., almond, citrus, grape, peach) and to seed (e.g., citrus). The importance of these methods of transmission in shade trees, however, is not known. Since disease development within populations of trees is most often random, direct tree-to-tree spread is unlikely. It is not known, however, how long urban forest trees remain asymptomatic following infection, thus other methods of transmission may, indeed, occur.
Mechanism of disease development
X. fastidiosa lives and multiples within the tracheary elements, tracheids, vessels, and intercellular spaces of xylem tissue (Figure 19). Compared to phloem, xylem fluid is nutritionally poor but does consist of amino, organic, and inorganic acids. These compounds, especially the amino acids glutamine and asparagine, are used by both the bacterium and the insect vector for growth. The quality and composition of xylem fluid varies both between hosts and within a host by season, time of day, and with the health and age of the host plant. This affects the feeding behavior of the vector; insects move from plant to plant to obtain the right combinations of nutrients needed for different stages of growth. Those insects that harbor
X. fastidiosa transmit the pathogen to new hosts as they search for suitable sources of nutrients.
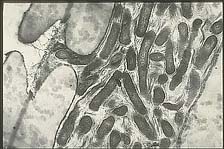
Figure 19a |
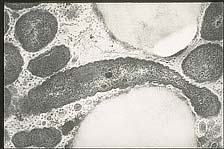
Figure 19b |
Cells of
X. fastidiosa attach to xylem vessel walls as well as to the foregut of insect vectors by producing biofilms (Figure 20). Bacterial cells aggregate together and are encased in a self-produced matrix of polysaccharide. This protects the cells and is thought to enhance pathogenicity. Terminal fimbriae (also called type IV pili) are important for biofilm formation. In addition, although the bacterium lacks flagella for motility, terminal fimbriae aid in a type of incremental movement called “twitching motility,” which enables bacterial cells to move against the xylem stream.

Figure 20a |

Figure 20b |
Symptoms of water stress (evident as leaf scorch) result from high populations of bacterial cells in xylem tissue as well as overproduction of defense compounds, such as pectins, and tyloses, produced by the host plant in response to infection. Embolisms (or air bubbles) eventually form and help to plug affected xylem vessels, leading to reduced xylem function and water stress. When prolonged, tissue available for photosynthesis is reduced and starch reserves are depleted, resulting in leaf scorching and premature senescence.
Populations of bacterial cells within xylem tissue fluctuate seasonally. In diseases where stunting is a primary symptom, such as phony peach disease and alfalfa dwarf, bacteria congregate in the roots. In hosts where leaf scorch is a primary symptom, such as BLS of shade trees, the pathogen resides above ground, and populations are greatest in the veins and petioles of symptomatic leaves. Most strains of
X. fastidiosa are sensitive to cold, and in shade trees, the pathogen is presumed to overwinter in more protected parts of the tree, such as the trunk or roots, and as the growing season progresses, populations increase and the pathogen moves acropetally to distal portions of the canopy. Indeed, most
Xylella-associated diseases occur in regions of North America where the winters are mild, such as the southeastern U.S. For example, the threat of Pierce’s disease is low where the average January temperature is less than 36 F. Strains that cause bacterial leaf scorch of urban forest trees, however, are more cold hardy and have been detected as far north New York and southern Ontario. These reports, however, are rare; the pathogen that affects oak has the most northern range (the disease is epidemic in certain regions of New Jersey) and may be more tolerant of cold than those than affect sycamore and elm, which are more prevalent in the mid-Atlantic states and south. Although cold hardiness of the pathogen may explain the geographic distribution of
Xylella-associated diseases, vector movement may also play a role. For bacterial leaf scorch of shade trees, these questions have yet to be answered.
Disease Management
Historically, management of the many diseases caused by
X. fastidiosa has encompassed various strategies such as reducing host stress, planting hosts not known to harbor the bacterium, and removing infected hosts, vectors, and alternative hosts. In shade trees, it is not known whether therapeutic pruning (removing infected branches as they become symptomatic) or removing infected trees stops the spread of the disease, both within a tree or within populations of trees. Indeed, the random incidence of BLS within a tree planting leads one to suspect that
X. fastidiosa also does not spread through root grafts, but this method of transmission has yet to be investigated. Management of BLS is made further difficult because it is not known how long hosts may remain asymptomatic prior to the first expression of symptoms.
The use of the antibiotic oxytetracycline to control
X. fastidiosa has been attempted on hosts such as grape, plum, and shade trees. Such trunk or root flare injections, however, provide only seasonal relief from symptoms, do not work well in trees with advanced disease, and must be repeated annually to be effective. Further studies are needed to determine if such antibiotic injections are effective or phytotoxic long-term or if repeated wounding during the injection process has a negative impact on the tree.
Management of the diseases caused by
X. fastidiosa through vector eradication has been attempted, but even with Pierce’s disease, where disease vectors are known, results are inconclusive. Trials evaluating the systemic insecticide imidacloprid are underway in infected shade trees. This compound is applied in a non-invasive manner to trees (as a soil injection) and is retained in the canopy for a 3-yr period. Ultimately, vector eradication may be useful only to prevent or reduce the rate of spread to adjacent plantings. More novel approaches to management of
X. fastidiosa in grapes, such as biological control or biofilm disruption, are currently under study, but use of such techniques for BLS of shade trees may not be feasible for years to come.
In summary, there are few cost-effective methods for the management of BLS in landscape plantings. Current management recommendations include:
Maintain plant vigor. The best management tool for this disease is to maintain tree vigor. The development of oak leaf scorch is enhanced by other diseases, insects, and environmental stresses such as drought. Bacterial leaf scorch may also predispose infected plants to other disease and insect problems.
Practice sanitation. Branches that have died due to oak leaf scorch should be routinely removed. Infected trees that are in a severe state of decline should also be removed.
Use tolerant plants. In areas where BLS occurs, avoid planting highly susceptible trees, and design new tree plantings with a diverse complement of tree species. Management of BLS in many regions of the eastern United States may ultimately depend on the identification of germplasm tolerant to the disease.
Significance of the Disease
History and Etiology
In 1892, Newton B. Pierce (Figure 21), state plant pathologist of California, examined grapevines (Vitis vinifera) with a scorch and decline of unknown cause (Table 6). At that time, the problem was known as California vine disease. Although the disease was eventually named for him, Pierce could neither isolate nor culture the causal agent, and he ended his career suspecting that a “minute microorganism” was involved. About the same period, peach growers in Georgia observed that many trees were stunted (“pony trees”) and the fruit they produced was undersized and unmarketable (in a disease charmingly called “phony peach disease”). It would be many years before scientists could prove that the causal agent of these two seemingly unrelated maladies was related.

Figure 21 |
Table 6. Discovery of the identity of
Xylella fastidiosa.
|
1890s | California vine disease (now known as Pierce’s disease) and phony peach disease observed and studied in different parts of the United States; As Newton B. Pierce states, a "minute microorganism" may be involved. |
1936 to
1959 | Root graft, budding, and/or insect transmission of Pierce’s disease, alfalfa dwarf, and elm scorch; causal agent considered to be a virus. |
|
1971 | Tetracycline antibiotic suppresses development of Pierce’s disease; causal agent now considered to be a mycoplasma-like organism (MLO). |
|
1973 | Electron microscopy reveals a "Rickettsia-like bacterium" in xylem tissues associated with Pierce’s disease and phony peach disease; elm leaf scorch still considered to be caused by a virus. |
|
1978 | Selective media were defined and a bacterium isolated from infected grapevine xylem tissues. |
|
1980 | A xylem-limited bacterium was associated with leaf scorch of elm. |
|
1987 |
Xylella fastidiosa described as a new bacterial species. |
|
2004 | New subspecies for
Xylella fastidiosa described based on pathogenicity, phylogenetic characteristics, and DNA relatedness. |
Clues to the etiology of what would become known as “Xylella-associated diseases” began to emerge in 1936 when both Pierce’s disease and alfalfa dwarf were transmitted through root grafts and by insects. Because of these properties, the causal agent was considered to be viral in origin until 1971, when antibiotics applied to grapevines with Pierce’s disease suppressed symptom development. Since viruses are not subject to control by antibiotics, the organism was reclassified as a mycoplasma-like organism (or MLO). In 1973, electron microscopy revealed an organism in the xylem vessels of infected grapevine leaves and peach roots that had a rippled cell wall and could not be cultured on standard culture media. This “rickettsia-like” bacterium (RLB) was subsequently found in many hosts, and the list of these “Xylella-associated diseases” grew.
Study of these diseases was significantly advanced in 1978 when a culture medium sufficient to isolate and grow the bacterium
in vitro was developed. In 1987 this organism was properly described as a bacterium closely related to the bacterial plant pathogen
Xanthomonas and was given the name
Xylella fastidiosa (“Xylella” because it grows in the xylem; “fastidiosa” because the organism is fastidious and difficult to grow in culture).
X. fastidiosa is one of several “xylem-limited bacteria.” More information on this type of pathogen can be found at:
http://publish.apsnet.org/edcenter/intropp/PathogenGroups/Pages/Fastidious.aspx
Today, diseases caused by
X. fastidiosa continue to increase in importance as entire crops such as grapes, citrus, and coffee, in certain locations are threatened. Since the late 1800s, wine grapes in California were protected from Pierce’s disease by planting grapevines at least 100 m from bodies of water where alternative host vegetation predominates. This strategy worked for over 100 years because the vectors of this disease were unable to traverse this distance from alternative host vegetation to the grape plantings. Recently, however, the glassy winged sharpshooter (H. coagulata), common in peach and plum orchards in the southeastern United States, was introduced to the grape-growing regions of California. This voracious feeder is a strong flier and has the potential to easily spread Pierce’s disease to valuable grape plantings. Many resources are now allocated for research to help prevent the potential devastation to the California wine industry from Pierce’s disease.
X. fastidiosa is of particular concern in South America, where plantings of citrus and coffee are affected by closely related, if not identical, strains. Coffee leaf scorch (or Requeima do Café) causes leaf scorch (as the name suggests) as well as premature leaf drop and shoot dieback. Symptoms of citrus variegated chlorosis (CVC) include small fruit with hard rinds that cannot be marketed. CVC has been of such importance that this was the first of various
X. fastidiosa strains to have its genome completely sequenced.
The incidence of BLS in shade trees is equally important from both economic and aesthetic points of view. For example, in some communities in New Jersey (Figure 22), BLS affects as many as 35% of street and landscape oaks. Current loss of value plus replacement costs for older trees affected by this disease is estimated at $8,000 per tree. Landowners and tree care professionals in these locations must plan for the loss of property values and high costs of replacement as shade trees in landscapes, wood lots, and golf courses affected by BLS decline and must be removed.
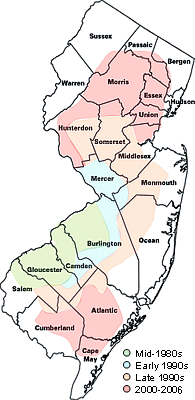
Figure 22 |
Selected References
Brlansky, R.H., L.W. Timmer, W.J. French, and R.E. McCoy. 1983. Colonization of the sharpshooter vectors,
Oncometopia nigricans and
Homalodisca coagulata, by xylem-limited bacteria. Phytopathology 73: 530-535.
Davis, M.J., A.H. Purcell, and S.V. Thomson. 1978. Pierce's disease of grapevines: Isolation of the causal bacterium. Science 199: 75-77.
Hammerschlag, R., J. Sherald, and S. Kostka. 1986. Shade tree leaf scorch. Journal of Arboriculture 12: 38-43.
Hearon, S.S., J.L. Sherald, and S.J. Kostka. 1980. Association of xylem-limited bacteria with elm, sycamore, and oak leaf scorch. Canadian Journal of Botany 58: 1986-1993.
Hewitt, W.B., B.R. Houston, N.W. Frazier, and J.H. Freitag. 1946. Leafhopper transmission of the virus causing Pierce's disease of grape and dwarf of alfalfa. Phytopathology 36: 117-128.
Hopkins, D.L. 1989.
Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annual Review of Phytopathology 27: 271-290.
Hopkins, D.L., and W.C. Adlerz. 1988. Natural hosts of
Xylella fastidiosa in Florida. Plant Disease 72: 429-431.
Marques, L.L. R.H. Ceri, G.P. Manfio, D.M. Reid, and M.E. Olson. 2002. Characterization of biofilm formation by
Xylella fastidiosa in vitro. Plant Disease 86: 633-638.
Meng, Y., Y. Li, C.D. Galvani, G. Hao, J.N. Turner, T.J. Burr, and H.C. Hoch. 2005. Upstream migration of
Xylella fastidiosa via pilus-driven twitching motility. Journal of Bacteriology 187: 5560-5567.
Purcell, A.H., A.H. Finlay, and D.L. McLean. 1979. Pierce's disease bacterium: Mechanism of transmission by leafhopper vectors. Science 206: 839-841.
Schaad, N.W., E. Postnikova, G. Lacy, M.B. Fatmi, and C.-J. Chang. 2004.
Xylella fastidiosa subspecies:
X. fastidiosa subsp.
piercei supsp. nov.,
X. fastidiosa subsp.
multiplex subsp. nov., and
X. fastidiosa spbsp.
pauca subsp. nov. Systematic and Applied Microbiology 27: 290-300.
Sherald, J.L. 2001.
Xylella fastidiosa, a bacterial pathogen of landscape trees. Pages 191-202 in Shade Tree Wilt Diseases, edited by C.L. Ash. American Phytopathological Society, St. Paul, MN.
Sherald, J.L., J.M. Wells, S.S. Hurtt, and S.J. Kostka. 1987. Association of fastidious, xylem-inhabiting bacteria with leaf scorch in red maple. Plant Disease 71: 930-933.
Simpson, A.J.G., et al. 2000. The genome sequence of the plant pathogen
Xylella fastidiosa. Nature 406: 151-157.
Wells, J.M., B.C. Raju, H.-Y. Hung, W.G. Weisburg, L. Mandelco-Paul, and D.J. Brenner. 1987.
Xylella fastidiosa gen. nov., sp. nov: Gram-negative, xylem-limited, fastidious plant bacteria related to
Xanthomonas spp. International Journal of Systematic Bacteriology 37:136-143.

