Common smut (Syn. boil smut, blister smut)
Ustilago maydis (Syn. Ustilago zeae)
Maize (Zea mays), teosinte (Zea mexicana)
Common smut of corn, caused by Ustilago maydis, is easily identified by tumor-like galls that form on actively growing host tissues and contain masses of dark, sooty teliospores. Throughout most of the world, common smut is considered to be a troublesome disease of corn, but in central Mexico, galls on ears of corn are considered an edible delicacy known as cuitlacoche (Syn. huitlacoche). In addition to the practical significance of causing a prevalent disease and being an edible fungus, U. maydis also has been used as a model organism to study a variety of interesting biological phenomena.

Common smut of corn caused by Ustilago maydis. (Courtesy J.K. Pataky)
Key words: boil smut, common smut, corn smut, cuitlacoche, huitlacoche, maize mushrooms, Ustilago maydis, Ustilago zeae
Symptoms and Signs
Ustilago maydis infects two hosts: maize (Zea mays) and teosinte (Zea mexicana). Like other smut diseases, common smut of corn derives its name from the sooty masses of teliospores found on infected host plants (Figure 2). The most obvious symptoms are tumor-like galls that vary in size from less than 1 cm to more than 30 cm (0.4-12 inch) in diameter. All meristematic tissues are susceptible to infection. Galls are found most frequently on ears, tassels, stalks, nodal shoots, and mid-ribs of leaves (Figures 3-6). Even though galls may form on many above-ground parts of the plant, infection is local (i.e., the host is not colonized systemically). Occasionally, galls develop beneath the soil surface when the apical meristem of a young plant is infected.

Figure 2 |

Figure 3 |

Figure 4 |

Figure 5 |

Figure 6 |
Smut galls consist of fungal and host tissues. Young galls are white, firm and covered with a semiglossy periderm. As galls begin to mature, interior tissue becomes semifleshy and streaks of black tissues occur as teliospores begin to form. With further maturation, galls become a mass of powdery teliospores and the periderm ruptures releasing the spores (Figures 2 and 3). Galls usually are obvious within 10 to 14 days after infection. On ears, galls mature about three weeks after ovaries are infected. Slightly discolored or disfigured kernels are obvious 3 to 6 days after infection (Figure 7). Small, white, firm galls develop 9 to 10 days after infection, and galls begin to enlarge resulting in a tumor-like growth 11-12 days after infection (Figure 8). Galls start to have a gray, silvery appearance as streaks of blackened tissues (teliospores) begin to form 14 to 15 days after infection (Figures 9a,b,c). About 70% of the gall tissue is blackened while galls retain a semi-fleshy, mushroom-like integrity 16 to 18 days after infection (Figures 10 a,b). By 21 to 23 days after infection, the periderm ruptures and galls become a sloppy, wet, mass of teliospores. With dehydration, galls become sooty, powdery masses of teliospores.

Figure 7 |

Figure 8 |

Figure 9a |

Figure 9b |
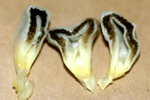
Figure 9c |

Figure 10a |

Figure 10b |
The number, size and location of galls depend on the age of plants at the time of infection. Leaf galls differ greatly in size but usually are small when compared to stalk and ear galls. Galls 20-30 cm (8-12 inches) in diameter are common on the stalk. Rudimentary ear shoots below the fertile ear are commonly infected. Galls can replace most of the tassel or individual florets depending on the time and severity of infection (Figure 5). Ear galls usually result from infection of individual ovaries. Commonly, a few kernels at the basal or tip ends of the ear are infected (Figure 11), although nearly every kernel on an ear may be replaced by smut galls if most ovaries are infected.
Symptoms of head smut of corn, caused by Sphacelotheca reiliana, are somewhat similar to common smut (Figure 12). Head smut occurs when maize plants are infected systemically by S. reiliana. Floral structures (tassels and ears) are converted to sori containing masses of powdery teliospores that resemble mature galls of common smut. Vascular bundles of the host usually are present as thread-like structures in sori of S. reiliana.

Figure 11 |

Figure 12 |
Pathogen Biology
Ustilago maydis (Syn. Ustilago zeae) is taxonomically grouped with the heterobasidiomycete fungi. The life cycle of U. maydis includes three distinct stages. Diploid teliospores are formed in galls on infected hosts and are the overwintering propagules of the fungus. They are spherical to ellipsoid, olive-brown to black in color and heavily echinulated, i.e., covered with tiny spines (Figure 13). When a teliospore germinates, it forms a septate promycelium, undergoes meiosis, and forms haploid sporidia (also called basidiospores) that usually have a single nucleus (Figure 14). The haploid sporidia are easily maintained in culture which many considered to be the saprophytic stage of the fungus. Sporidial cultures can be propagated on many different media where they display considerable variation in morphology, color, size and growth (Figure 15). The non-infectious sporidia bud in a yeast-like manner (Figure 16). Under appropriate environmental conditions, genetically compatible sporidia mate and form dikaryotic infection hyphae; cells in this third stage are pathogenic and can infect corn and teosinte.
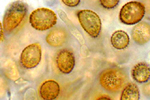
Figure 13 |

Figure 14 |
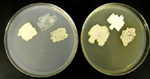
Figure 15 |
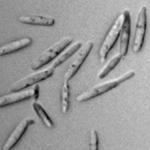
Figure 16 |
Additional details about the biology of Ustilago maydis
Mating and infection are under genetic control of two different loci, a and b. Historically, the a locus has been called the mating type locus and the b locus has been called the pathogenicity locus. In order for U. maydis to infect, mating partners (i.e., haploid sporidia that form diakaryotic hyphae) must have different alleles at both the a and b loci (e.g., a sporidium with a1b2 alleles mates with one having a2b1 alleles). Alleles from both loci have been cloned and sequenced. The a locus has two alleles, a1 and a2, and encodes components of a pheromone response pathway which mediates formation of conjugation tubes and cell fusion. When nutrients are limiting, haploid sporidia that have unlike alleles at the a locus form slender filaments that grow towards each other following pheromone gradients. Ultimately, the filaments fuse. Multiple alleles occur at the b locus which encodes DNA-binding proteins with regulatory functions. If the dikaryon that results from filament fusion has two nuclei with unlike alleles at the b locus, a vigorous infection filament forms (Figure 17). The infection filament responds to an appropriate surface of the host by forming an appressorium that allows the fungus to penetrate the host (Figure 18). Appressoria form only on living cells that are rapidly elongating. Appressoria have not been induced in vitro. Mating and penetration occur within a 12-18 hr period on plants artificially inoculated with compatible sporidia.
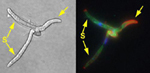
Figure 17 |

Figure 18 |
In the first several days of infection, infected tissues may appear slightly chlorotic, but there are few macroscopic signs of infection. Microscopic examination reveals that in these early stages, the U. maydis dikaryon behaves as a biotroph. The hyphae grow mainly intracellularly, but both host and fungal plasma membranes remain intact. The plant cells undergo normal cytoplasmic streaming even though they are invaded by the hyphae (Figure 19). Regular haustoria are not observed, but the presence of an electron-dense matrix material in between the host and fungal walls is a likely site of nutrient transfer (Figure 20.). The exact nature of the nutrients transferred is not known, but most U. maydis strains that are amino acid auxotrophs are non-pathogenic, suggesting that a simple carbohydrate may be the primary nutrient used by the fungus.

File is a .MOV file
and will require a movie
viewer on your browser to play.
You can download Quicktime
if you need a viewer. |
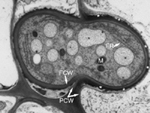
Figure 20 |
Microscopic alterations such as abnormal enlargement of parenchyma cells may be seen as early as 24 hrs after inoculation. Tumors resulting from abnormal growth of host cells begin to develop as early as a week after infection. Tumors initially result from abnormal cell division and enlargement in host tissues, and fungal hyphae are relatively rare. Host cells remain alive during early stages of tumor formation, continuing to grow and divide, but the normal development of tissues is disrupted. For example, the regular pattern of venation is lost, and parenchyma cells may become extremely large. As the tumors enlarge, hyphae begin to proliferate in between the host cells rather than growing through them (Figure 21). Karyogamy (i.e., fusion of nuclei that occurs during sexual reproduction) appears to occur in early stages of tumor enlargement, and is followed by rapid hyphal proliferation in between host cells. At this stage, the hyphal walls become swollen and gelatinous, and the protoplasts lose their cylindrical shape and become spherical. By processes that are poorly understood, the spore initials enlarge and develop the pigmented, ornamented teliospore walls, which may be seen emerging from the remnants of the gelatinized hyphae (Figure 22). The fungus completes its life cycle each time teliospores form on an infected host.
Figure 21 |

Figure 22 |
Disease Cycle and Epidemiology
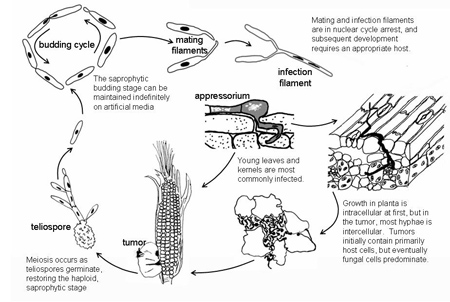 Disease Cycle
Disease CycleEpidemiology
Near the end of the 19th century, Brefeld worked out many of the details of the disease cycle of common smut which coincides with the life cycle of U. maydis. The fungus overwinters as diploid teliospores in crop debris or soil. Christensen estimated that a single smut gall of medium size may contain more than 200 billion spores (Figure 23). Teliospores can remain viable for several years. Teliospores can be disseminated directly by wind or splashing rain, or they can germinate and undergo meiosis to form haploid sporidia which also can be disseminated by wind or splashing rain. Sporidia bud in a yeast-like manner, mate and form dikaryotic infection hyphae. Infection hyphae of compatible mating types (i.e., different alleles at the a and b loci) penetrate and infect the host. All infection is local. Any above-ground plant part can be infected, particularly young actively growing meristematic tissues. The fungus also may grow down stigmas (ear silks) and into developing ovaries (kernels), resulting in ear galls. Each gall on an infected ear is the result of infection of an individual ovary. Disfigured tissue which develops into galls may be noticeable within days after infection. Galls form within a week after infection and enlarge for up to 3 weeks after infection. About 1 to 2 weeks after infection, hyphae in galls begin to gelatinize, and hyphal cells become round and form cell walls of teliospores. As galls mature and dehydrate, the periderm ruptures releasing teliospores.

Figure 23 |
Teliospores produced in galls of infected tissues are probably unimportant as inocula in the growing season in which they are produced. However, they become the overwintering inocula that initiate infection on subsequent crops. There appears to be sufficient inoculum in nature to result in as much common smut as host and environmental factors will allow. In some instances, common smut has occurred in fields that were newly broken and have never been planted with corn previously.
There is no general agreement on weather conditions that are most favorable for common smut although most reports indicate that common smut is prevalent following rainy, humid weather. There also are reports linking increased infection rates with wounding of plants, as may occur during detasseling. Galls on leaves and stalks of seedlings often are observed following strong thunderstorms with heavy winds, especially when plants are injured by blowing soil. Obviously, germination and dissemination of teliospores and/or sporidia would be favored by wind and rain. However, anything that induces rapid localized cell division and elongation, such as wounding, could increase susceptibility to smut infection. Factors that reduce the production of pollen or inhibit pollination also increase the occurrence of ear galls of common smut because ovaries are protected from infection by U. maydis soon after they are fertilized, mostly likely because silks attached to fertilized ovaries die and are thus no longer susceptible to infection. For example, hot, dry, drought-like conditions often cause asynchronous pollen production and silk emergence which results in poor pollination. Ear galls of common smut may be prevalent if U. maydis is readily disseminated to stigmas of unfertilized ovaries during or immediately following these hot, dry conditions. Thus, some people associate the occurrence of ear galls with droughts although the droughts probably affect the prevalence of ear galls primarily by increasing the number of unpollinated ovaries with rapidly growing silks.
Common Smut of Corn Disease Management
Management
Many methods of controlling common smut have been recommended or evaluated, including crop rotation, sanitation, seed treatments, application of foliar fungicides, modification of fertility, and biological controls. In spite of these frequently mentioned control tactics, host resistance is the only practical method of managing common smut in areas where U. maydis is prevalent. Nevertheless, no corn line is immune from infection by U. maydis.
Resistance
The first significant advance in the control of common smut occurred when the first generation of corn hybrids were developed from inbred lines that where selected for lower incidence of smut. Although resistance to common smut has been studied for nearly 100 years, specific mechanisms of resistance and genes that condition resistance have not been documented clearly.
Early corn breeders observed that varieties of flint corn tended to be more smut-susceptible than varieties of dent corn. In 1918, just a decade after George Shull and Edward East independently discovered heterosis (hybrid vigor) in corn, Donald Jones demonstrated that inbred lines of corn developed by him and East at the Connecticut Agriculture Experiment Station differed strikingly in the number of smut infected plants. During the next ten years, early corn breeders such as H. K. Hayes, R. J. Garber and F. R. Immer reported that self-fertilization of open-pollinated maize varieties rapidly led to the isolation of inbred lines with noticeably different reactions to common smut. Not only did inbred lines differ in the incidence of infected plants, susceptible lines could be differentiated by the plant parts on which galls formed. For example, some inbreds were uniformly infected on lower nodes of the stalk while others had a high incidence of ear or tassel galls. The rapidity with which selfed lines became homogeneous for smut reactions led Immer and J.J. Christensen to speculate that “a relatively small number of genetic factors are involved in determining smut reactions,” and that “composite crosses [of smut-resistant lines] will undoubtedly yield smut-resistant lines in a large percentage of cases.” Fortunately, the latter of these two hypotheses proved to be correct.
In 1928, Marion Griffiths, a USDA plant pathologist, demonstrated that selfed lines of corn and crosses between them that were resistant to natural infection by U. maydis in the field were very susceptible when inoculated. None of the resistant lines were immune from infection when Griffiths used a needle to inject sporidial suspensions of U. maydis into young plant tissues. She hypothesized that “resistance or susceptibility is largely a matter of relative accessibility of the susceptible [plant] parts to the invading organism.” Other researchers soon began to corroborate Griffith’s observations by demonstrating that protection of susceptible host tissues by non-susceptible tissues played an important role in smut resistance. After observing a strong association between smut susceptibility and the open or closed condition of apical buds (i.e., tightness of leaf whorls), A. H. Eddins concluded that “open buds hold the inoculum and allow [spore] suspensions to seep down the plant and come in contact with young, actively growing and susceptible tissues.”
J.M. Walter and others observed that injury to leaf and stalk tissues or injury resulting from detasseling plants increased the incidence of common smut although the smut galls did not occur on the injured tissues themselves, but instead, occurred on meristematic tissues in close proximity to the injured tissues. After evaluating 200 inbred lines grown under smut-epidemic conditions for 10 years and crosses between those lines and genetic linkage testers, M. M. Hoover concluded that smut reactions probably were conditioned by two sets of genetic factors: one controlling physiological behavior and one concerned with gross morphology of the plant.
By the early 1940s when double-cross hybrids were beginning to be grown on a significant amount of the corn acreage in the U.S., corn breeders had substantially improved smut resistance simply by selecting against smut susceptibility in breeding populations and among inbred lines. When G. H. Stringfield and D. H. Bowman compared the incidence of smut on 340 open-pollinated varieties and 1,052 corn hybrids in 48 experiments throughout Ohio from 1933 to 1938, they observed that 75% of the hybrids had less smut than the open-pollinated varieties (Figure 24). Hybrids derived from one of twelve inbreds lines that Stringfield considered to be smut resistant had less than a third as much smut as the open-pollinated cultivars. This type of progress, resulting from selection against smut susceptibility without knowledge of the specific resistance genes involved or the mechanisms of resistance, has continued through today. Most modern field corn hybrids grown in the U.S. have adequate levels of resistance to natural infection to avoid catastrophic smut epidemics. Nevertheless, no hybrid is known to be immune to infection when injected with sporidial suspensions, and major genes responsible for resistance have not been identified definitively in spite of recent evaluations of mapping populations for quantitative trait loci for smut resistance.
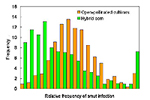
Figure 24 |
Sweet corn hybrids, whose pedigrees often include New England flint germplasm, tend to be more susceptible to common smut than field corn. This may be due to differences in the genetic backgrounds of field corn and sweet corn and/or differences in the diligence with which susceptible lines have been eliminated from field corn and sweet corn breeding programs. Ear galls are of greatest economical importance in sweet corn. Similar to the conclusions made by breeders and pathologist in the 1920s and 1930s about the association between gross morphology and reaction of corn lines to smut, resistance to ear galls could be morphological or physiological. K. M. Snetselaar and co-workers recently demonstrated that ovaries (kernels) are protected from U. maydis infection when an abscission layer forms at the base of corn silks soon after fertilization (i.e., pollination). The rapidity with which abscission layers form following fertilization, the rate of pollen germ tube growth, synchrony of silk emergence and pollen production, and the height of ear placement (assuming that inoculum is dispersed from the soil) are just a few of the morphological factors that could allow corn plants to escape ear infection and appear to be ‘field resistant’.
Eradication and Exclusion
Although Ustilago maydis occurs throughout the world where corn is grown, the occurrence of common smut is localized in Australia where the disease is known as maize boil smut. For most of the 20th century, Australians attempted to exclude U. maydis from areas where it did not occur. Infected crops were destroyed to eradicate occurrences of U. maydis, and quarantines were enforced to exclude new introductions.
In 1911, when common smut was first reported in Bathurst, New South Wales, Australia, the infected crop was destroyed and corn was not planted on that farm for 10 years. The disease remained localized to that area, reoccurring sporadically between 1935 and 1940. Affected crops were destroyed and a quarantine enacted in 1940 prohibited corn from being grown in that area for 25 years. Also, areas free of the disease could only be planted with maize seed produced from crops free of U. maydis and treated with an approved fungicide. This was considered to be a successful case of eradication and exclusion because additional occurrences of common smut were not reported until 1982 when the disease was found throughout the North Coast district of New South Wales and southeast Queensland (Figure 25). By 1996, the disease spread to other areas of this region and crossed the Great Dividing Range where it spread southward to areas west of Sydney. The disease subsequently was reported from areas in southern New South Wales where corn is grown under irrigation and from at least one location in northern Victoria. According to corn seed companies in Australia, boil smut has continued spread in areas near Bathurst, Dubbo, and the Murrimbigee Irrigation district.
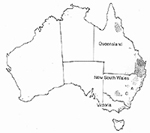
Figure 25 |
Significance
Economic significance
Economic losses from common smut have been relatively minor in field corn grown in the United States since the early 1940s when hybrids were widely adopted and levels of host resistance were improved dramatically. Nevertheless, yield can be reduced occasionally when smut is prevalent and severe. Losses are greatest when apical meristems of very young seedlings are infected which often results in plant death or barren plants. Ear galls also can be economically important in field corn depending on the severity of infection (i.e., the number of kernels per ear replaced by smut galls). Economically significant outbreaks of common smut in field corn usually are associated with severe thunderstorms that occur soon after seedlings emerge or plant stresses (e.g., drought) that result in asynchronous timing of silk emergence and pollen production. Prior to the development of hybrid corn, losses due to common smut were estimated to range up to 10% annually because the disease sometimes was quite prevalent. For example, incidence of infected plants was 6 to 18% when corn fields near Ames, Iowa were surveyed from 1930 to 1934. Other occurrences of over 75% incidence of smut infection of open-pollinated cultivars were reported occasionally prior to the development of hybrids.
Common smut is of greater economic consequence in sweet corn than in field corn. When sweet corn is grown for fresh market (e.g., roadside stands, farmers’ markets, retail stores), ears with a single smut gall usually are not marketable because of “cosmetic injury” (i.e., lack of consumer acceptance). Hence, even a very low incidence of common smut can result in substantial losses. When sweet corn is grown for processing (i.e., canning, freezing), fields with moderate amounts of ear galls often are not harvested. In fresh market and processing sweet corn crops, galls on plant tissues other than ears can cause secondary losses if the crop is mechanically harvested because husk leaves of ears become covered with teliospores (Figure 26). In processing crops, additional costs are incurred to remove teliospores that contaminate husk leaves. In fresh market corn, some consumers will not accept sooty, black teliospores on husk leaves even though ears are not damaged.

Figure 26 |
In areas of the world where open-pollinated corn cultivars are grown or where hybrid corn is developed from smut-susceptible germplasm, common smut continues to be problematic. For example, incidence of smut infection was greater than 50% in 1976 in several areas of Germany where hybrids derived from European flint corn were prevalent.
Model organism for basic biological studies
Ustilago maydis has been used by investigators as a model organism to study a variety of interesting biological phenomena, including fungal mating type, fungal dimorphism, plant-pathogen interactions, and genetic recombination and repair. It has an intriguing feature of a life cycle that has both biotrophic and saprophytic stages. Other features that make the fungus a particularly attractive experimental organism include: a) the saprophytic stage is haploid, and thus easily manipulated genetically, b) meiotic progeny are easily analyzed, c) the entire life cycle can be completed in young plants in about 3 weeks, and d) diploid strains can be artificially constructed in the laboratory. Only a few examples of these studies can be mentioned here.
In the 1960s and 70s, R. Holliday built on the work of previous workers when he carefully developed standard media and techniques for working with U. maydis. He also produced an impressive array of mutants of various kinds. Some of these mutants were especially susceptible to additional mutations, because they were defective in their ability to repair their DNA. Holliday used these mutants to develop a model for how homologous recombination can occur between paired DNA molecules. The “Holliday structure” and Holliday’s model of genetic recombination are described in virtually every college genetics textbook. The work on recombination has continued to bear fruit, most notably in the lab of W. Holloman. Women with inherited mutations in a human tumor suppressor gene known as BRCA2 are more likely to develop breast and ovarian cancer, compared to women without the mutations. Workers in Holloman’s lab found the homolog of BRCA2 in U. maydis, and were able to study how the products of this tumor suppressor gene are involved in making repairs to damaged DNA.
Mating compatibility systems in filamentous basidiomycetes are characterized by multiple alleles at two genetic loci; strains with unlike alleles at both loci are said to be compatible, and can mate and begin sexual development. The multiplicity of possible combinations of alleles has fascinated scientists studying mushrooms and other basidiomycetes for many years, but the molecular interactions were not understood until recently. Mating compatibility in U. maydis was well-described by Christensen and previous workers, and it is easy to assay haploid isolates for mating compatibility using a charcoal agar assay first described by Day and Anagnostakis (Figure 27). In the late 1980s and early 1990s, S. Leong, R. Kahmann, and co-workers took advantage of earlier work, and the fact that U. maydis was easily grown as a haploid yeast in liquid culture when they developed and used a genetic transformation system that allowed them to clone and describe the mating compatibility genes in U. maydis. The presence of a pheromone-receptor combination at one locus coupled with a transcriptional regulator at the other locus, first elucidated in U. maydis, is the common theme in basidiomycete mating systems.

Figure 27 |
Most recently, the entire genome of U. maydis has been sequenced as part of an international collaboration. The genome and its annotation are publicly available at the Ustilago maydis database maintained by the Munich Information Center for Protein Sequences (MIPS) and online at http://mips.gsf.de/projects/fungi/ustilago.html. The availability of the genomic sequence and ancillary tools are beginning to bear fruit for scientists interested in host-pathogen interactions.
Cuitlacoche: edible galls of Ustilago maydis
Cuitlacoche (or huitlacoche) is the native Mexican name given to young, edible galls that form when ears of corn are infected by Ustilago maydis. In central Mexico, cuitlacoche is a highly prized delicacy that has been eaten since Pre-Columbian times. Traditional maize growers gather and market cuitlacoche following natural infection. About 400 to 500 tons of cuitlacoche are sold annually during July and August at markets in Mexico City (Figure 28a-d). More than 100 tons are processed by companies that sell the specialty mushroom canned or lyophilized (Figure 29a,b).

Figure 28a |

Figure 28b |

Figure 28c |

Figure 28d |

Figure 29a |

Figure 29b |
Concurrent with an expanding market in the U. S. for other types of specialty mushrooms such as Pleurotus (oyster), Lentinula (shiitake), Flammulina (enoki) and Morchella (morel), epicureans in North America increasingly view cuitlacoche as a gourmet fungus that is part of a growing market for haute Mexican cuisine. Cuitlacoche is served in soups, appetizers and entrees at many fashionable Mexican restaurants in major metropolitan areas in the United States, such as Topolobampo in Chicago and Rosa Mexicano in New York City and Washington DC (Figure 30a-c). Recipes for cuitlacoche are available on the internet and in gourmet Mexican cookbooks, such as Rick Bayless’s Mexican Kitchen and Diana Kennedy’s The Art of Mexican Cooking. Canned cuitlacoche is sold on the internet and sometimes is referred to as “maize mushrooms” or “Mexican truffles”. Fresh or frozen cuitlacoche occasionally is available at farmers’ markets or from local suppliers in the U.S. (Figure 31a-c).

Figure 30a |

Figure 30b |

Figure 30c |

Figure 31a |

Figure 31b |

Figure 31c |
Methods to cultivate cuitlacoche as a cash crop and various other aspects of cuitlacoche production systems have been studied in the U.S. and Mexico during the past 15 years. Essential aspects of efficient cultivation of cuitlacoche include: efficient methods of inoculation, rates of gall enlargement and teliospore formation, optimal times for inoculation and harvest, production practices that optimize infection, gall size, and yield, and, possibly most important, proper post-harvest handling and marketing.
Selected References
Banuett, F. 1995. Genetics of Ustilago maydis, a fungal pathogen that induces tumors in maize. Annual Review of Genetics 29:179-208.
Bayless, R., D.G. Bayless, and J.M. Brownsen. 1996. Rick Bayless’s Mexican Kitchen. Scribner, New York. 448 pp.
Christensen, J.J. 1963. Corn smut caused by Ustilago maydis. American Phytopathological Society Monograph 2. 41 pp.
Day, P.R., and S.L. Anagnostakis. 1971. Corn smut dikaryon in culture. Nature: New Biology 231:19-20.
Kahmann, R., and J. Kämper. 2004. Tansley Review: Ustilago maydis: How its biology relates to pathogenic development. New Phytologist 164:30.
Kennedy, D. 1989. The Art of Mexican Cooking: Traditional Mexican Cooking for Aficionados. Bantam Books. New York. 526 pp.
Pataky, J. K. and M. A. Chandler. 2003. Production of huitlacoche, Ustilago maydis: timing inoculation and controlling pollination. Mycologia 95:1261-1270.
Snetselaar, K.M., M. Carfioli, and K.A. Cordisco. 2001. Pollination can protect maize ovaries from infection by Ustilago maydis, the corn smut fungus. Canadian Journal of Botany 79:1390-1399.
Wang, J., D.W. Holden, and S.A. Leong. 1988. Gene transfer system for the phytopathogenic fungus Ustilago maydis. Proceedings of the National Academy of Science 85:865-869.
Yang, H., Q. Li, J. Fan, W. Holloman, and N. Pavletich. 2005. The BRCA2 homologue Brh2 nucleates RAD51 filament formation at a dsDNA-ssDNA junction. Nature 433:653-657.
Links to Websites
The MIPS Ustilago maydis Genome DataBase
http://mips.helmholtz-muenchen.de/genre/proj/ustilago
Saint Joseph’s University Ustilago Research Page
http://www.sju.edu/biology/ksweb/ustilago/intro.html
Broad Institute Ustilago maydis data base
http://www.broad.mit.edu/annotation/fungi/ustilago_maydis/
Various recipes for cuitlacoche
http://www.sweetcorn.uiuc.edu/Common-smut/Recipes.htm

