Westphal, A., T.S. Abney, L.J. Xing & G.E. Shaner. 2008. Síndrome de la muerte súbita de la soja. Trans. Silvina L. Giammaria 2008. The Plant Health Instructor. DOI:10.1094/PHI-I-2009-0217-01
Síndrome de la muerte súbita (SMS) de la soja
Norteamérica: Fusarium virguliforme (Aoki et al., 2003). Sinónimo: F. solani (Mart.) Sacc. f.sp. glycines; Sudamérica: F. brasiliense, F. cuneirostrum, F. tucumaniae y F. virguliforme (Aoki et al., 2005).
Soja (o soya) (Glycine max)
Autores
Andreas Westphal1, T. Scott Abney2, Lijuan Xing1 y Gregory Shaner1
1 Purdue University, y 2 USDA-ARS, West Lafayette, IN.
Traductora: Silvina L. Giammaria, Estación Experimental Agroindustrial “Obispo Colombres”, Las Talitas, Tucumán, Argentina.
Traducción revisada por Eduardo Gallego & José Sánchez, Área de Botánica, Universidad de Almería, Almería, España, y Walter Gutierrez, USDA-APHIS, Raleigh, NC.
El síndrome de la muerte súbita (SMS) de la soja (o soya) fue descubierto en Arkansas en 1971, y desde entonces su presencia ha sido confirmada en la mayoría de las áreas sojeras de EE.UU. El SMS es una enfermedad fúngica, pero también se presenta en combinación con el nematodo del quiste de la soja (NQS, Heterodera glycines), constituyendo un complejo patogénico. El SMS es una de las enfermedades causadas por patógenos de suelo más devastadoras de la soja en EE.UU. Cuando la enfermedad se da en presencia del NQS los síntomas aparecen más temprano y son más severos. Los síntomas de la enfermedad son más notorios después de la floración.
Síntomas y signos
Generalmente, el síndrome de la muerte súbita (SMS) de la soja no se detecta en el follaje de las plantas hasta después del inicio de la floración. Bajo circunstancias atípicas, las plantas más jóvenes pueden presentar algunos síntomas. Cuando se hacen prospecciones de la enfermedad, resulta útil comparar las plantas afectadas con las sanas del mismo campo.

Figura 1a |

Figura 1b |

Figura 1c |
Follaje y tallos
Los síntomas iniciales del SMS son un moteado clorótico difuso y un arrugamiento de las hojas. Más adelante, el tejido foliar amarillea entre las nervaduras principales, luego muere y empardece. Esta clorosis y necrosis internerval es típica del SMS, pero estos síntomas no son diagnósticos por sí solos porque pueden confundirse con los síntomas foliares de otras enfermedades. Poco después, los folíolos mueren y se marchitan. En casos severos, los folíolos se caen, dejando los pecíolos adheridos al tallo. Para el diagnóstico, el cuello (o corona) y la raíz principal deben abrirse en dos. El tejido cortical de una planta con el SMS exhibirá unas rayas que van desde el crema al marrón claro, mientras que el de una sana será blanco. Sin embargo, la médula (la porción central del tallo) de una planta infectada permanecerá blanca o crema clara.

Figura 2a |
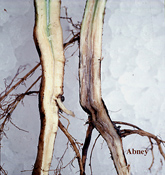
Figura 2b |

Figura 2c |
Raíces
Al extraerse una planta con síntomas severos de SMS, el sistema radical de esta última carecerá de vigor comparado al de una planta sana. Puede haber también podredumbre radical. Si las plantas se recolectan cuando el suelo está húmedo, pueden observarse pequeñas manchas azules claras en la superficie de la raíz principal al ras del suelo. Éstas son masas de esporas azules de los hongos que causan el SMS. A medida que la superficie radical se seca, el color azul desaparece, pero estas masas de esporas azules, junto con los otros síntomas mencionados anteriormente, son unos buenos indicadores de diagnóstico del SMS.
Distinción del SMS de otras enfermedades similares
La podredumbre marrón del tallo, el cancro del tallo, la podredumbre carbonosa y la quemadura química pueden producir síntomas foliares similares a los del SMS. Sin embargo, en estas enfermedades, los folíolos muertos tienden a permanecer adheridos a los pecíolos. La podredumbre marrón del tallo difiere de los síntomas del SMS porque la médula se torna oscura y hay una decoloración insignificante de la corteza (o córtex). El cancro del tallo suele asociarse con unos tapetes de esporas (en inglés, spore mats) dentro del tallo que se ven como estrías reticuladas longitudinales. La podredumbre carbonosa adquiere su nombre a partir del desarrollo de microesclerocios dentro del tallo y que se asemejan a una fina capa de carbonilla. Cuando los síntomas foliares los causa la quemadura (o quemado) química no se encuentran signos de ningún patógeno y las raíces de las plantas afectadas están sanas.
Biología del Patógeno
El SMS es causado por hongos de suelo ubicados dentro de un grupo (agrupamiento cladístico 2) del complejo de especies de Fusarium solani (Aoki et al., 2003; Aoki et al., 2005). En América del Norte, Fusarium virguliforme (Aoki et al., 2003), anteriormente Fusarium solani f. sp. glycines (Roy, 1997), es el agente causal. En Sudamérica, F. brasiliense, F. cuneirostrum, F. tucumaniae, y F. virguliforme causan síntomas del SMS. En inoculaciones en invernadero, F. cuneirostrum y F. phaseoli, agentes causales de la podredumbre radical del poroto (o frijol), también son capaces de causar síntomas del SMS en la soja (Aoki et al., 2005). Los hongos de este complejo taxonómico crecen lentamente en los medios de cultivo y son difíciles de aislar de las plantas enfermas. Una vez que se obtiene un cultivo puro, los patógenos causantes del SMS se distinguen de otras especies de Fusarium que puedan infectar raíces de soja en base a características culturales y a la presencia de las esporas azules. Se considera que, en América del Norte, el patógeno del SMS es de reproducción clonal y asexual, aunque experimentos recientes de apareamientos revelaron la existencia de un teleomorfo (Covert et al., 2007). En cruzamientos específicos bajo condiciones de laboratorio, uno de los patógenos del SMS en Sudamérica, F. tucumaniae, formó peritecios fértiles (estructuras sexuales de reproducción). Basado en el Código Internacional de Nomenclatura Botánica de Viena (Art. 59.7), se retuvo el nombre del estado anamorfo del hongo (Covert et al., 2007). Fusarium virguliforme, el patógeno del SMS en América del Norte, no formó cruzamientos fértiles consigo mismo o con F. tucumaniae; solamente se produjeron estructuras sexuales interfértiles en algunos de estos últimos cruzamientos (Covert et al., 2007). Estos resultados sustentan la hipótesis de que en América del Norte el patógeno del SMS es clonal y que las especies causantes de esta enfermedad son diferentes en América del Norte y del Sur, respectivamente.
Probablemente, entre los diferentes ciclos del cultivo de soja el patógeno del SMS sobrevive como clamidosporas (o clamidósporas) en el rastrojo o libres en el suelo (figura 3a). Las clamidosporas, de paredes gruesas, se desarrollan en el suelo y sobre las raíces de soja durante el ciclo de la enfermedad y de ahí en adelante. Pueden soportar amplias fluctuaciones en la temperatura del suelo, incluso bajo el punto de congelación, y resistir la desecación. A medida que el suelo se calienta en la primavera, las clamidosporas cercanas a las raíces de soja reciben el estímulo para germinar e infectarlas.
Aunque durante el verano el patógeno puede producir esporas (macroconidios, Figura 3b) sobre la superficie de la raíz principal, estas esporas sólo se dispersan a distancias cortas y dentro de la misma estación de crecimiento. A lo largo de los años, el flujo del agua y ciertas prácticas agrícolas que impliquen el movimiento de suelo favorecerán el desplazamiento de las esporas a distancias largas dentro del mismo campo o entre varios.
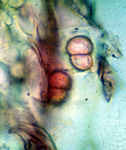
Figura 3a |
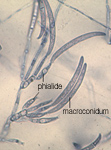
Figura 3b |
Ciclo de la Enfermedad y Epidemiología
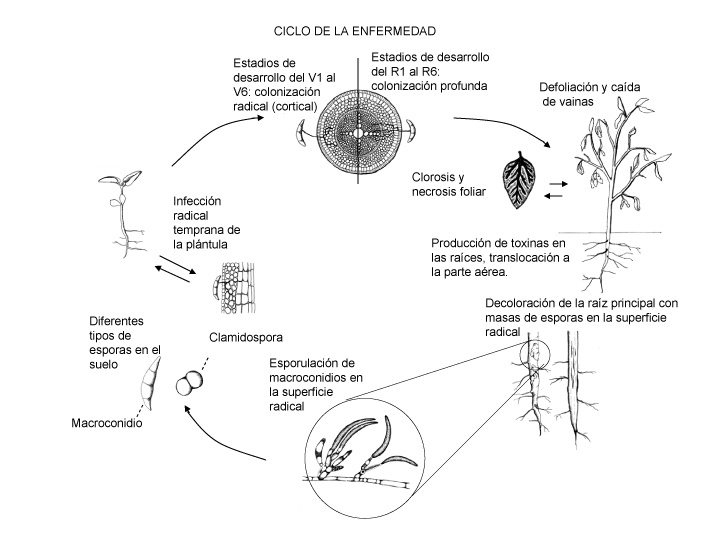
El hongo puede infectar las raíces de plántulas de soja apenas después de la siembra, pero los síntomas aéreos del SMS raramente aparecen antes de mediados de julio en la región del medio-oeste de EE.UU., momento en el cual las plantas alcanzan los estadios reproductivos (N.T.: en el hemisferio sur, este estadio tendría lugar en torno a enero). En este momento, el hongo penetra en los tejidos vasculares de la planta (Roy et al., 1997). El hongo produce toxinas en las raíces que se translocan hacia las hojas. Frecuentemente, los síntomas aparecen por primera vez después de las fuertes lluvias que se dan en los estadios reproductivos; la elevada humedad edáfica incrementa la severidad de la enfermedad (Xing & Westphal, 2006). Las toxinas son las que causan los síntomas foliares ya que el hongo no invade los tallos mas allá de unos pocos centímetros por encima del nivel del suelo (Roy et al., 1997).
Los primeros síntomas visibles del SMS son un amarillamiento y una defoliación de las hojas superiores. Apenas aparecen en el campo, los síntomas suelen estar confinados en pequeñas áreas (o rodales) o franjas, generalmente en las áreas más húmedas o compactadas (Figura 4), como las cabeceras del cultivo (en inglés, turn rows). Después de dos o tres semanas, las áreas afectadas se agrandan y algunas plantas de otras áreas del campo pueden mostrar síntomas.
El grado de pérdidas de rendimiento que causa el SMS depende de la severidad y el momento de la expresión de la enfermedad en relación al desarrollo de la planta con respecto a los componentes del rendimiento. Si la enfermedad se da al principio de los estadios reproductivos, puede haber aborto de flores y de vainas jóvenes. Cuando la enfermedad es tardía, la planta producirá menos semillas por vaina o semillas más pequeñas. Mientras más temprana sea la enfermedad, mayor será la reducción del rendimiento. Dado que el hongo del SMS persiste en el suelo por períodos largos, el área afectada será mayor cada año, hasta llegar a un punto donde todo el campo estará afectado. No se ha desarrollado ningún umbral de inóculo del patógeno, pero una hipótesis es que el SMS aparece siempre y cuando el patógeno esté presente y las condiciones sean conducentes para la enfermedad. El SMS es más severo cuando el nematodo del quiste de la soja (NQS, Heterodera glycines) también está presente en el campo y el cultivar es susceptible a ambos patógenos (Xing & Westphal, 2006). Tanto F. virguliforme como el NQS están muy difundidos en los campos de soja de EE.UU. La asociación tan estrecha de ambos patógenos se evidencia también en el hecho de que el patógeno del SMS puede aislarse a partir de los quistes del NQS (Roy et al., 1997). No se entiende bien cuáles son las implicancias de esto para la epidemiología de la enfermedad. Las prácticas productivas que mantienen bajos los niveles del NQS podrían reducir el riesgo de incidencia del SMS, pero se necesitan más investigaciones al respecto para poder entender la naturaleza de la interacción entre estos dos patógenos. Si el campo estuviese infestado con el patógeno del SMS, se debería hacer una prospección para ver si el NQS está presente. Si es así, debería implementarse una estrategia de manejo (o gestión) conjunto para ambas enfermedades.

Figura 4 |
Manejo (o gestión) de la enfermedad
Las opciones de manejo del SMS son limitadas. A pesar de que existen cultivares de soja menos susceptibles al SMS, no hay cultivares altamente resistentes a la enfermedad (Njiti et al., 2002). La aplicación de fungicidas en el surco durante la siembra o como tratamiento de semillas sólo tienen un efecto limitado en la reducción de la enfermedad. La aplicación de fungicidas foliares no tiene ningún efecto en la supresión del SMS debido a que la infección fúngica está restringida al sistema radical y los fungicidas, normalmente, no se mueven hacia abajo por lo cual no alcanzan el sitio de infección. Varias prácticas de manejo pueden reducir el riesgo de daño por el SMS, aunque ninguna de ellas puede prevenir la enfermedad.
Fecha de siembra
Las siembras tempranas predisponen a la soja para la infección. En suelos húmedos y frescos, las plántulas de soja son vulnerables a la infección del patógeno del SMS. Si las condiciones primaverales favorecen el rápido crecimiento de la soja, y no hay lluvias que saturen el suelo durante los estadios reproductivos tempranos, entonces el riesgo del SMS puede ser menor, aunque el hongo esté presente en el suelo. Los campos sin historial del SMS deberían ser los primeros en sembrarse; los campos donde sí es un problema deberían ser los últimos.
Labranza
Los suelos compactados impiden la percolación del agua y restringen el crecimiento radical. Una fuerte lluvia en el momento en que la soja ha alcanzado los estadios reproductivos saturará las áreas compactadas, lo cual promueve el desarrollo del SMS. El riesgo del SMS puede reducirse si se corrigen los problemas de compactación y permeabilidad. El suelo responde de forma diferente a los cambios impuestos por la intensidad de los sistemas de labranza; el arado, cincelado o similares perturbaciones drásticas del suelo afectan fuertemente el drenaje, la posición de los rastrojos, y la composición microbiana del suelo (Aon, 2001; Kladivko, 2001). No sorprende que los informes respecto a los efectos de la labranza en el desarrollo del SMS sean contradictorios. En algunos suelos, la siembra directa (o labranza cero) puede ayudar a reducir la severidad del SMS en comparación con las labranzas con el arado o la combinación de arado y cincel (Seyb et al., 2007; Abney, sin publicar). En otros suelos, la labranza intensiva reduce los niveles del SMS, probablemente porque mantiene el adecuado movimiento vertical de agua en comparación con la labranza reducida (Vick et al., 2006). El mejor drenaje posible y las condiciones de desarrollo radical más propicias pueden facilitarse de diversas maneras—en algunos suelos, puede requerir el uso de una labranza intensiva, mientras que en otros tipos de suelo, la labranza reducida puede ser la más apropiada.
Rotación
La rotación de cultivos puede reducir el riesgo del SMS (Rupe et al., 1997), pero la combinación de maíz-soja en rotación anual, muy común en el cinturón maicero, no reduce la incidencia ni la severidad de la enfermedad (Westphal, sin publicar). Severas epifitias se dieron aún después de muchos años de un continuo cultivo de maíz. La rotación de cultivos reduce el potencial de inóculo de otros patógenos de la soja, pero el cambio a rotaciones anuales de maíz y soja (comparado con rotaciones más largas que incluyen a otras gramíneas y forrajeras) no es efectivo para reducir el riesgo del SMS. Estudios realizados en la Universidad de Purdue encontraron que después de la rotación con maíz, las raíces de soja no estaban visualmente más sanas que las que provenían del monocultivo de soja (Xing & Westphal, sin publicar). Cuando se siembra maíz, la población de patógenos edáficos puede declinar hasta cierto punto, pero no lo suficiente como para reducir la presión de la enfermedad cuando se siembre soja al año siguiente. Aunque una rotación de dos años puede mantener las densidades poblacionales del NQS por debajo de los umbrales si la densidad poblacional inicial es baja, dicha rotación es demasiado corta para reducir el riesgo del SMS.
Cultivares resistentes
Los mejoradores de soja se esfuerzan en desarrollar cultivares resistentes al SMS, pero el progreso ha sido lento. Se desarrollaron algunos métodos de campo e invernadero, que usaban altas tasas artificiales de inóculo sobre granos de sorgo y una selección cuidadosa de los regímenes de riego (Hartman et al., 1997; De Farrias et al., 2006). Aunque la mayoría de las empresas de semillas han eliminado de sus inventarios los cultivares altamente susceptibles, no existen actualmente cultivares altamente resistentes. Dado que las compañías de semillas continuamente introducen al mercado nuevos cultivares y retiran los más viejos, es esencial contar con información precisa de la reacción de los nuevos cultivares al SMS.
Bajo estas condiciones especiales de manejo, debe evitarse la siembra de cultivares de soja altamente susceptibles en campos con alto riesgo del SMS. Para el manejo del SMS y otras enfermedades edáficas, es fundamental mantener un registro de campo de cuándo y dónde se presentaron las mismas. Los dispositivos portátiles de GPS pueden ser de utilidad para cumplir este objetivo, pero un buen plano del campo con las áreas problemáticas puede ser suficiente. Los campos severamente afectados con SMS deberían identificarse para sembrarlos más tarde y realizar en ellos operaciones que mejoren la permeabilidad del agua, incluyendo prácticas de labranza que corrijan la compactación o el drenaje. Finalmente, deberían sembrarse cultivares con algún grado de resistencia.
Importancia
El síndrome de la muerte súbita (SMS) es una de las enfermedades más importantes de la soja en América del Norte y del Sur (Aoki et al., 2005; Wrather & Koenning, 2006; Wrather et al., 2003). El SMS fue informado por primera vez en América del Sur a comienzos de la década de 1990 (Roy et al., 1997). En EE.UU., el SMS se descubrió por primera vez en Arkansas en 1971. Desde entonces, la enfermedad se dispersó ampliamente y es la causa actual de la mayor parte de las pérdidas de rendimiento que se dan en la región norte-centro. El SMS es más grave en siembras tempranas en suelos húmedos y frescos, cuando las fuertes lluvias estivales saturan el suelo y cuando el NQS está presente.
Agradecimientos
Este manuscrito está publicado con el número 2007-18145 del Programa de Investigación Agrícola de la Universidad de Purdue (Purdue University Agricultural Research Program). El trabajo artístico de K. A. Frank (ciclo de la enfermedad) fue enormemente valioso.
Referencias seleccionadas
Aoki, T., K. O’Donnell, Y. Homma, and A.R. Lattanzi. 2003. Sudden-death syndrome of soybean is caused by two morphologically and phylogenetically distinct species within the Fusarium solani species complex-F. virguliforme in North America and F. tucumaniae in South America. Mycologia 95:660-684.
Aoki, T., K. O'Donell, and M.M. Scandiani. 2005. Sudden death syndrome of soybean in South America is caused by four species of Fusarium: Fusarium brasiliense sp. nov., F cuneirostrum sp. nov., F. tucumaniae, and F. virguliforme. Mycoscience 46:162-183. Online: doi:10.1007/s10267-005-0235-y.
Aon, M.A., D.E. Sarena, J.L. Burgos, S. Cortassa. 2001. (Micro)biological, chemical and physical properties of soils subjected to conventional or no-tillage management, and assessment of their quality status. Soil and Tillage Research 60:173-186. Online: doi:10.1016/s0167-1987(01)00190-8.
Covert, S.F., T. Aoki, K. O'Donnell, D. Starkey, A. Holliday, D.M. Geiser, F. Cheung, C. Town, A. Strom, J. Juba, M. Scandiani, X.B. Yang. 2007. Sexual reproduction in the soybean sudden death syndrome pathogen Fusarium tucumaniae. Fungal Genetics and Biology 44:799-807. On-line: doi:10.1016/j.fgb.2006.12.009.
De Farias Neto, A., G.L. Hartman, W.L. Pedersen, S. Li, G. Bollero, and B.W. Diers. 2006. Irrigation and inoculation treatments that increase the severity of soybean sudden death syndrome in the field. Crop Science 46:2547-2554. Online: doi: 10.2135/cropsci2006.02.0129.
Hartman, G.L., Y.H. Huang, R.L. Nelson, and G.R. Noel. 1997. Germplasm evaluation of Glycine max for resistance to Fusarium solani, the causal organism of sudden death syndrome. Plant Disease 81:515-518.
Kladivko, E. 2001. Tillage systems and soil ecology. Soil and Tillage Research 61:61-76. Online: doi:10.1016/s0167-1987(01)00179-9.
Njiti, V.N., K. Meksem, M.J. Iqbal, J.E. Johnson, M.A. Kassem, K.F. Zobrist, V.Y. Kilo, and D.A. Lightfoot. 2002. Common loci underlie field resistance to soybean sudden death syndrome in Forrest, Pyramid, Essex, and Douglas. Theoretical and Applied Genetics 104:294-300. Online: doi: 10.1007/s001220100682.
Roy, K.W. 1997. Fusarium solani on soybean roots: nomenclature of the causal agent of sudden death syndrome and identity and relevance of F. solani form B. Plant Disease 81:259-266.
Roy, K.W., D.E. Hershman, J.C. Rupe, and T.S. Abney. 1997. Sudden death syndrome of soybean. Plant Disease 81:1100-1111.
Rupe, J.C., R.T. Robbins, and E.E. Gbur, Jr. 1997. Effect of crop rotation on soil population densities of Fusarium solani and Heterodera glycines and on the development of sudden death syndrome of soybean. Crop Protection 16:575-580.
Seyb, A., L.J. Xing, T.J. Vyn, J. Seo, T.S. Abney, and A. Westphal. 2007. Tillage system effects on sudden death syndrome, Heterodera glycines, and soybean yield in a Mollisol. Phytopathology 97:S106 (abstract).
Vick, C.M., J.P. Bond, S.K. Chong, and J.S. Russin. 2006. Response of soybean sudden death syndrome to tillage and cultivar. Canadian Journal of Plant Pathology 28:77-83.
Wrather, J.A., and S.R. Koenning. 2006. Estimates of disease effects on soybean yields in the United States 2003 to 2005. Journal of Nematology 38:173-180.
Wrather, J.A., S.R. Koenning, and T.R. Anderson. 2003. Effect of diseases on soybean yields in the United States and Ontario (1999-2002). Plant Health Progress. Online: doi:10.1094/PHP-2003-0325-01-RV.
Xing, L.J., and A. Westphal. 2006. Interaction of Fusarium solani f. sp. glycines and Heterodera glycines in sudden death syndrome of soybean. Phytopathology 96:763-770. Online: doi:1094/PHYTO-96-0763.

