Take-all root rot
Gaeumannomyces graminis
small grains (wheat, barley, oats), weedy and wild grasses, and bentgrasses (creeping, colonial, velvet).

|
Take-all is an important root disease of cereal crops and bentgrasses throughout the world. The study of this disease has contributed much to our understanding of biological control of root pathogens.
Symptoms and Signs
Field symptoms
On small grains, take-all first becomes apparent near the time when the seed head emerges. The leaves are yellow and plants may be stunted. The disease usually occurs in circular patches (Figure 2) although it also can be fairly uniform throughout a field (Figure 3, right three-quarters). Perhaps the most diagnostic field symptom is prematurely-ripe tillers ("whiteheads") (Figure 4).

Figure 2 |

Figure 3 |

Figure 4 |
On closely mowed bentgrass turf, take-all results in the formation of roughly circular dead patches ranging from a few centimeters (inches) to more than a meter (3 feet) in diameter (Figure 5). Patch margins often are bronze to yellow-orange. Patches may reappear in the same location in successive years.

Figure 5 |

Figure 6 |

Figure 7 |
Root Symptoms
Take-all is identified by the dark brown to black rotten roots or stolons. Initially, roots have small black lesions that expand and later coalesce (Figure 6). Heavily rotted roots are very brittle and much of the root system remains in the soil when plants are pulled up(Figure 7). When the leaf sheaths are pulled away from the stem, a shiny black discoloration of the basal stem is highly diagnostic for take-all (Figure 7). Under a microscope, dark brown "runner hyphae" (or ectotrophic growth, Figures 8 and 9) and mycelial mats (Figure 10) on roots or stolon surfaces are easily seen and are also helpful in diagnosing this root disease.

Figure 8 |

Figure 9 |

Figure 10 |
Pathogen Biology
The fungus persists from season to season in crop debris. The mycelium infects roots of living hosts (Figures 6 & 8) and as the root dies, the fungus saprophytically colonizes the dying tissue (Figure 11). Therefore, the fungus is almost exclusively soil-borne. Infested, soil-borne fragments can be transported by wind, water, animals, and farm or turf machinery. The fungus is a relatively poor saprophyte and does not compete well with the native soil microflora. It is also sensitive to inactivation by heat when soils are heated by solar energy. Thus, it is most prolific when host roots are plentiful and when there are relatively short saprophytic periods (weeks or a few months) between the availability of susceptible roots.
Figure 6 |
Figure 8 |

Figure 11 |
Gaeumannomyces graminis is the name given to the sexual or "perfect" stage of this Ascomycete fungus. The fungus produces dark, flask-shaped fruiting bodies called perithecia (Figure 12). Each perithecium (Figure 13) contains many asci (sacs). Each ascus (Figure 14, left) contains eight long, thin spores called ascospores (Figure 14, right). The sexual stage of the fungus is not important to the development of the disease.

Figure 12 |
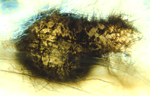
Figure 13 |
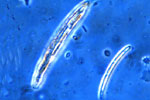
Figure 14 |
Strains or varieties of the Gaeumannomyces graminis fungus are associated with different grass hosts. For example, the variety tritici (G. graminis var. tritici) causes root rot on wheat, the variety avenae affects roots of oats and bentgrasses, and the variety graminis causes a sheath rot of rice and is associated with a root decline of bermudagrass, centipedegrass, and St. Augustinegrass in the southeastern United States.
Biological control
An interesting phenomenon called "take-all decline," associated with wheat monoculture, has been described in many parts of the world. When this happens, the disease declines to an insignificant level after several years of being severe. Root-colonizing bacteria, that are antagonistic to the take-all fungus, build up in the soil to make it suppressive to take-all. An important aspect of suppression is the production of antibiotics by the bacteria that inhibit the growth of the fungus on the root. Figure 15 shows the inhibition (clear zone) of the take-all fungus by three colonies of antagonistic bacteria. Take-all decline is one of the classic examples of "biological control" of a plant disease and has been extensively studied.
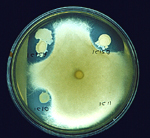
Figure 15 |
Disease Cycle and Epidemiology
During time periods when no living hosts are present, the fungus survives as mycelium in soil-borne host residue that was infected the previous growing season (Figure 16). It also can survive parasitically on roots of volunteer hosts, perennial hosts, or grassy weeds. The sexual stage (ascospores) is not important.

Figure 16 |
When roots of a susceptible plant grow near to the infested residue, the mycelium colonizes the roots and begins the infection process. "Runner hyphae" on the root surface (Figure 17) produce infections (Figure 18) at numerous locations along the roots.
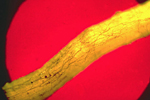
Figure 17 |
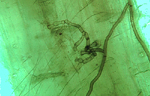
Figure 18 |
Infections progress into the stele of the root (Figure 19) and move up and down the root axis resulting in root death. When numerous roots are killed (Figure 6), or the fungus reaches the crown area and rots it, the plant dies.
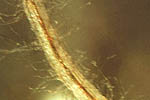
Figure 19 |
Figure 6 |
After plant death, the fungus survives saprophytically in the plant tissues that it colonized during its parasitic phase, completing the cycle (Figure 16).
Epidemiology
Take-all on small grains occurs worldwide. Fall-sown small grains are more severely damaged than spring-sown ones. The disease is important only where these hosts are grown in a monoculture (that is, where crop rotation is not practiced). It is usually more severe under reduced-tillage regimes, soils that are between pH 6.0 and 8.5, moist soil conditions, and soil temperatures between 5 and 15°C (40-60° F). Although it is usually most severe in coarse-textured soils where there is good aeration, take-all can occur in heavier soils.
Take-all also occurs on bentgrass throughout North America, western Europe, Australia, and New Zealand. It is most common when bentgrass is planted into recently cleared land or soil that has been fumigated. It may be particularly severe on recently established, sand-based golf course putting greens. It is believed that the increased severity of take-all in newly-seeded bentgrass and in sand-based greens is due to the limited populations of antagonistic bacteria. As in cereal crops, infection and colonization of roots is favored by cool, wet conditions in fall and spring, although disease symptoms may be even more apparent following hot, dry conditions in summer.
Disease Management
Crop rotation: Rotations to non-host species or a weed-free fallow are the most effective method of controlling take-all of small grains (Figure 3, extreme left). Because the fungus is a poor saprophyticcompetitor, even a one-year break from production gives almost complete control. During the rotation or fallow period, grassy weeds that can harbor the fungus (Figure 20) should be controlled.
Figure 3 |

Figure 20 |
Cultural practices:
Tillage: Although there are conflicting reports from Europe, conventional tillage in the U.S., such as with a moldboard plow (Figure 21), usually reduces take-all over reduced-tillage methods that leave more residue intact on the soil surface. There are three main reasons for less take-all with tillage in the U.S. First, the infested debris is broken into smaller fragments and exposed to more rapid decay, thereby weakening the fungus. Second, burying the crop residue (Figure 22, right) eliminates its soil shading effect and as a result the high soil temperatures of summer kill the fungus. Third, tillage kills grassy weeds and volunteer plants that harbor the fungus.
Weed and volunteer control: The take-all fungus can survive on the roots of grassy weeds and volunteer cereals. When these weeds or volunteer are killed by glyphosate, the defenses of the plant are turned off, because of the mode of action of this herbicide. The take-all fungus can then increase on the dying plant and "green-bridge" to the newly planted crop. Therefore, it is recommended that growers kill out weeds and volunteers at least 2-3 weeks before planting.

Figure 21 |

Figure 22 |
Soil drainage and compaction: Good soil drainage produces an environment less conducive to take-all root rot. Therefore, selection of well-drained sites and soil drainage methods should be practiced where possible. Aerification (removal of soil cores) is also effective in reducing disease severity in bentgrass turf areas.
Soil pH: Soil pH can have a profound effect on severity of the disease. Soils below pH=6.0 limit the disease. On the other hand, liming to raise soil pH frequently results in severe take-all. Therefore, crop rotation or a fallow period should be coupled with lime applications.
Seed bed: A firm seed bed for small grains is preferable to a loose one. A firm seed bed reduces the spread of the pathogen along the roots.
Planting date: Delayed planting in the fall reduces take-all by prolonging the time that the fungus is in the saprophytic phase. Because the pathogen is a poor saprophyte, lengthening the saprophytic period weakens the fungus thus exposing it to further microbial activity.
Fertilizers: As a general rule, ammoniacal (e.g. ammonium sulfate) forms of nitrogen reduce the disease over nitrate forms by reducing root surface pH. Some organic soil amendments also have been shown to reduce take-all. Recently, foliar applications of manganese sulfate have been shown to reduce take-all severity on bentgrass turf.
Irrigation: For small grain crops in areas where irrigation is available (Figure 23), applications of water should be as infrequent and deep as possible. Allowing the soil to dry out between irrigations is important to slow down the rate of root colonization and subsequent colonization of the crown.

Figure 23 |

Figure 24 |
Maintaining biological control: A natural suppressiveness to take-all can build up in the soil in a monoculture. Fields that show "take-all decline," or those in long-term continuous production of hosts, can show suppressiveness. Unfortunately, rotation to non-grass crops usually eliminates the suppressiveness of a soil and any decision concerning rotations should take this into account. Biological control research using seed treatments with suppressive bacteria (Figure 24, left) on small grains or applying biological agents to established bentgrass has shown promise. Commercial applications of this technology may be available in the future.
Resistant cultivars: Although there are small grain and bentgrass cultivars that have a slight level of resistance to take-all, no highly-resistant cultivars are available. Some level of resistance has been identified in wild grass species that may be transferable into cultivated species. One mechanism for increased resistance is the formation of a barrier or "apposition" by the host (Figure 25) in an attempt to stop penetration by the take-all fungus.
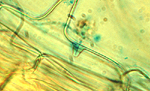
Figure 25 |

Figure 26 |
Fungicides: Foliar fungicides have no effect on this root disease of small grains. Two new seed treatment fungicides, silthiofam and fluquinconazole, are commercially available in Europe and the UK, and can reduce and control take-all in certain situations (Figure 26, right) However, these are not registered in the US.
Applications of certain sterol-inhibiting (DMI) and strobilurin fungicides are effective in suppressing take-all symptoms on bentgrass turf and are usually part of an overall management strategy. One or two fungicide applications are usually made in the fall for several consecutive years.
Historical Significance
Take-all has been recognized as a problem on small grains for about 150 years since first being described in Australia in 1852. The wheat farmers in Australia gave the disease its name around 1870 because of the severity of its attack and the devastation that it caused their crops. The actual cause of the disease, however, was unknown for almost 50 years with many strange hypotheses advanced including "clouds of small flies." Finally in 1890, researchers in France correctly ascribed the malady to a root-rot fungus. The original name they gave to the fungus (Ophiobolus graminis) was used for many decades until the current name (Gaeumannomyces graminis) was shown to be the correct one.
Take-all is a disease of temperate regions, although it can also occur at higher elevations in tropical areas. Therefore, it occurs worldwide wherever small grains are grown. Because the fungus is a natural inhabitant of the roots of wild grasses, the pathogen has a worldwide distribution. When soils are put into small grain production, the disease can become problematic. Little progress has been made in the use of host resistance and chemicals to control take-all on small grains. Therefore, the disease continues to be an important problem.
Selected References
Asher, M.J.C. and P.J. Shipton. 1981. Biology and Control of Take-all. Academic Press, London.
Bateman, G.L., R.J. Gutteridge, and J.F. Jenkyn. 2004. Take-all and grain yields in sequences of winter wheat crops testing fluquinconazole seed treatment applied in different combinations of years. Ann. Appl. Biol. 145:317-330
Bockus, W.W. 1983. Effects of fall infection by Gaeumannomyces graminis var. tritici and triadimenol seed treatment on severity of take-all in winter wheat. Phytopathology 73:540-543.
Clarke, B.B. and A.B. Gould. 1993. Turfgrass Patch Diseases Caused by Ectotrophic Root-Infecting Fungi. APS Press, St. Paul. MN.
Cook, R.J. 1981. The influence of rotation crops on take-all decline phenomenon. Phytopathology 71:189-192.
Cook, R.J. 1994. Problems and progress in the biological control of wheat take-all. Plant Pathology 43:429-437.
Dernoeden, P. H. and N.R. O'Neill. 1983. Occurrence of Gaeumannomyces patch disease in Maryland and growth and pathogenicity of the causal agent. Plant Dis. 67:528-532.
Hornby, D. 1998. Take-all Disease of Cereals: A Regional Perspective. CAB International. 384 pp.
Huber, D.M. and T.S. McCay-Buis. 1993. A multiple component analysis of the take-all disease of cereals. Plant Dis. 77:437-447.
Raaijmakers, J.M. and D.M. Weller. 1998. Natural plant protection by 2,4- diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Molecular Plant-Microbe Interactions 11:144-152.
Schumann, G.L. and J.D. MacDonald. 1997. Turfgrass Diseases: Diagnosis and Management. CD ROM. APS Press, St. Paul, MN.
Smiley, R. W., P. H. Dernoeden, and B. B. Clarke. 1992. Compendium of Turfgrass Diseases. 2nd ed. APS Press, St. Paul, MN.
Weller, D. M., J.M. Raaijmakers, B. B. McSpadden Gardener, and L.S. Thomashow. 2002. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathology 40: 309-348.

