El cancro del tallo por Phomopsis es una de las principales enfermedades que limitan el rendimiento del girasol (Helianthus annuus L.) en todo el mundo (Harveson et al. 2016). La enfermedad se describió por primera vez en 1980 en la región de Vojvodina (en la antigua Yugoslavia) y el hongo causal fue identificado como
Diaporthe helianthi (sin.
Phomopsis helianthi) (Muntañola-Cvetkovic et al. 1985). Después de este primer reporte, el cancro del tallo por Phomopsis se documentó en los años 80 y 90 en varios países productores de girasol, tales como Hungría (Nemeth et al., 1981), Bulgaria (Mihailova 1984), Estados Unidos (Hajdu et al. 1984; Herr et al. 1983; Yang et al. 1984), Francia (Lamarque y Perny 1985; Regnault 1985), Ucrania, Moldavia (Bogdanova et al. 1986), y Rusia (Scripka et al. 1993). En todos estos países, se asumió que
D. helianthi era el único agente causal del cancro del tallo por Phomopsis, aunque varios investigadores sospecharon que podía ser causada por más de una especie de
Diaporthe (Aćimović y Štraser 1982; Herr et al. 1983; Yang y Gulya 1984).
En 2010 se produjo una epidemia de cancro del tallo por Phomopsis en Minnesota, Dakota del Norte y Dakota del Sur, estados en los que se concentra más del 75% de la producción de girasol de los Estados Unidos. Utilizando el análisis de secuencias de ADN y la morfología, se identificó una especie no descripta antes,
D. gulyae, como causante del cancro del tallo por Phomopsis sumándose a
D. helianthi. Un experimento en invernadero que comparó la virulencia de los aislamientos de
D. helianthi y
D. gulyae indicó que ambos fueron agresivos sobre el girasol, aunque con niveles significativamente diferentes (Mathew et al. 2015b). Un año más tarde, en Australia, donde
D. helianthi es una especie exótica, se informó un brote de cancro del tallo por Phomopsis que afectó a lotes comerciales de girasol, identificándose que los patógenos responsables fueron
D. gulyae y dos especies no descriptas anteriormente:
D. kongii y
D. kochmanii (sin.
D. sojae). Los experimentos de patogenicidad en invernadero sobre girasol comparando los aislamientos de
D. gulyae,
D. kongii y
D. kochmanii mostraron que
D. gulyae fue más agresivo que los otros dos patógenos (Thompson et al. 2011). En 2015, también en Australia, se describieron cinco nuevas especies patógenas en girasol (Thompson et al. 2015) y, más recientemente, se informó a
D. novem como patógeno (Thompson et al. 2018). También en 2015, se confirmó que
D. stewartii causaba cancro del tallo por Phomopsis en el estado de Minnesota (Olson et al. 2017). Además de las especies de
Diaporthe descritas anteriormente, investigaciones en curso en Australia, Argentina y los Estados Unidos han revelado al menos quince especies de
Diaporthe, tanto patógenas como no patógenas, asociadas con el cancro del tallo en girasol (F. Mathew et al., inédito; S. Thompson et al., inédito; Zambelli et al., 2021).
Signos y síntomas

Figura 1: Infección de la hoja por cancro del tallo por Phomopsis
|

Figura 2: Infección por Phomopsis que progresa desde la hoja hacia el pecíolo
|

Figura 3: Lesiones características del tallo de color marrón claro centradas en los nudos debido a una infección temprana. Las lesiones crecerán y se oscurecerán a medida que la enfermedad avanza
|

Figura 4: Daño de la médula detrás de la lesión del tallo causada por
D. gulyae
|

Figura 5: Vuelco de plantas de girasol debido a la infección por Phomopsis
|

Figura 6: Lesiones del tallo características del tallo negro por Phoma
|

Figura 7: Lesiones del tallo características de la podredumbre del tallo medio por Sclerotinia
|
Figura 8: Conidios asexuales alfa (ovalados) y beta (filiformes)
|
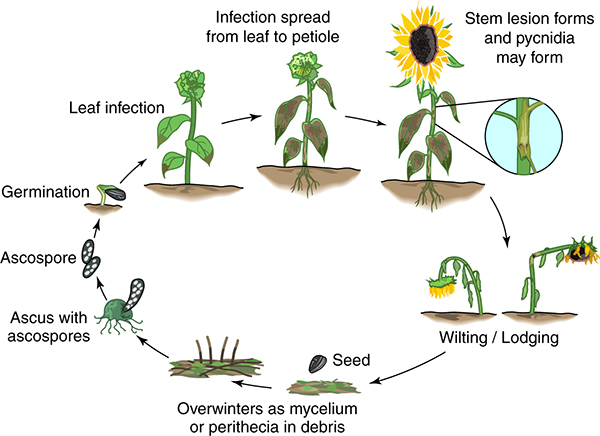
Figura 9: Ciclo de enfermedad del cancro del tallo por Phomopsis del girasol
|

Figura 10: Híbrido de girasol susceptible que presenta una infección severa por Phomopsis con vuelco de plantas
|
La infección del girasol por
D. helianthi (y posiblemente por
D. gulyae) se inicia durante las etapas de crecimiento vegetativo de las plantas. Los síntomas incluyen grandes manchas marrones de forma irregular en las hojas formando grandes parches de tejido muerto que se extienden desde los bordes. El tejido que rodea las venas afectadas puede ser clorótico, pudiendo luego pasar a necrótico y eventualmente provocar la muerte prematura de la hoja (Figura 1). La infección se propaga desde la hoja hacia el pecíolo, que se torna de color marrón oscuro (Figura 2) pasando luego al tallo donde se desarrolla una lesión necrótica de color marrón claro a oscuro en el nudo. A medida que la infección progresa, las lesiones aumentan de longitud (15 a 20 cm) las que en condiciones favorables pueden ceñir el tallo (Figura 3). A medida que la infección avanza, el hongo degrada el tejido de la médula debajo de la epidermis (Figura 4) lo que hace que el tallo se vuelva hueco y propenso al quebrado (Figura 5). Desde que se produce la infección de la hoja hasta que lesiona el tallo, pueden transcurrir entre 25 y 30 días.
Los síntomas del cancro del tallo por Phomopsis pueden confundirse fácilmente con los producidos por otros patógenos del tallo de girasol. Por ejemplo, algunas especies de
Diaporthe pueden producir una lesión en la planta de girasol similar a
Sclerotinia sclerotiorum (podredumbre del tallo por Sclerotinia) y especies de Phoma (tallo negro de Phoma). Sin embargo, a diferencia del cancro del tallo por Phomopsis, las lesiones del tallo negro por Phoma suelen ser más pequeñas, brillantes, de color negro, a menudo en forma de escudo (Figura 6), y generalmente no causan degradación de la médula. Las lesiones de la podredumbre del tallo medio por Sclerotinia pueden parecerse a las del cancro del tallo por Phomopsis, sin embargo las lesiones de Sclerotinia son de color tostado y tienen esclerocios negros que se forman dentro del tallo, los que a veces extruyen hacia el exterior a medida que la planta envejece (Figura 7) (Gulya et al. 1997).
Biología del patógeno
Durante el invierno
Diaporthe helianthi se mantiene en la superficie del suelo como peritecio sobre los rastrojos de cultivos infectados (Masirevic y Gulya 1992). En cuanto a
D. gulyae, las etapas periteciales del hongo rara vez se han observado en condiciones naturales (S. Thompson et al., inédito). Aun así, la diferenciación de la ascogonia en peritecio es un proceso lento que ocurre de forma desigual durante los meses de primavera. Los peritecios de
D. helianthi se forman sobre tejido cortical y son de forma globular a esférica, de color amarillo a negro, con un diámetro de 400 µm y cuellos largos que sobresalen (350 a 600 µm). Cuando los peritecios maduran en primavera, se producen numerosos ascos que son de forma globular o cilíndrica con tamaños que van desde 47,5 a 57,5 µm de largo x 7,5 a 12,0 µm de ancho. Los ascos maduran a una temperatura óptima de 25 °C (en un rango de 15 a 30 °C) y en condiciones húmedas y temperaturas inferiores a 10 °C liberan ascosporas a unos 3 mm por encima de la superficie de los rastrojos (Androsova et al. 2008). A veces, puede haber una masa blanca o una gota mucosa que contiene ascosporas producidas en el apéndice del rostrum del peritecio (Maric et al. 1982; Yakutkin 1988). Las ascosporas de
D. helianthi son bicelulares, incoloras, uniseptadas, elipsoidales y su tamaño varía entre 12,5 a 14,5 µm de largo x 3,2 a 4,5 µm de ancho (Muntañola-Cvetkovic et al. 1989). Las ascosporas de
D. helianthi son producidas por los peritecios durante aproximadamente 17 días (Li et al. 1985).
Durante el desarrollo de la enfermedad por
D. helianthi y
D. gulyae, los picnidios (de 170 a 3000 µm de diámetro) pueden ser producidos en pleno verano. Los picnidios son de color marrón oscuro, globulares, con picos ocasionales ostiolados y están incrustados en el tejido del hospedador. Cuando maduran, los picnidios producen alfa conidios y/o beta conidios en gotitas de color ámbar. Los alfa conidios de
D. helianthi y
D. gulyae son hialinos, de forma fusoide a elipsoide y con un tamaño que varía de 5,5 a 10,0 µm de largo x 1,5 a 3,0 µm de ancho (Figura 8). Los beta conidios de
D. helianthi son hialinos, filiformes y su tamaño varía entre 17,0 a 42,0 µm de largo x 0,5 a 2,0 µm de ancho (Figura 8) mientras que los de
D. gulyae no han sido observados
in vitro (Mathew et al. 2015b; Muntañola-Cvetkovic et al. 1989; Thompson et al.2015, 2011). El diámetro de los picnidios y la relación longitud / ancho de los conidios son variables entre las especies de
Diaporthe aisladas que causan enfermedades en el girasol.
Ciclo de la enfermedad y epidemiología
El ciclo de la enfermedad comienza con la producción y liberación de ascosporas de los peritecios que maduran sobre los rastrojos (Figura 9). Las ascosporas son arrastradas por el viento o salpicadas por la lluvia sobre las hojas medias o inferiores de las plantas de girasol durante la etapa de crecimiento. Estas esporas infectan los márgenes de las hojas más viejas de las plantas de girasol a través de gotitas de gutación en donde se acumula la humedad. Las lesiones iniciales se desarrollan cuando la humedad relativa en el canopeo supera el 90% durante 36 horas continuas (Debaeke et al. 2017). Sin embargo, en promedio, las lesiones foliares se vuelven visibles a los 20 a 25 días después de que las ascosporas causan la infección a temperaturas dentro del rango de 20 a 25 °C (Pinochet 1995). Además de la temperatura, la lluvia a partir de la diferenciación del botón floral (estadio fenológico R1; Berglund 2007) hasta el inicio de la floración (estadio fenológico R5) del girasol es el factor determinante para producir una infección foliar efectiva.
Después de que las hojas se infectan, los micelios colonizan los espacios intercelulares, las nervaduras secundarias y la nervadura central para luego avanzar por el pecíolo e invadir el tallo sistémicamente, en donde se formará una lesión alargada de color marrón tostado que se centra en las axilas (Muntañola-Cvetkovic et al. 1991). Las lesiones del tallo generalmente no se observan en las plantas de girasol hasta el momento de la floración (estadio R5). A medida que se coloniza el tallo, la hifa avanza a través del sistema vascular y se disemina a través del colénquima cubriendo la corteza, dando lugar a la formación de picnidios (Muntañola-Cvetkovic et al. 1989). Las pérdidas de rendimiento ocurren cuando los tejidos del xilema y el floema son invadidos, dejando el sistema vascular obstruido por las hifas fúngicas. Posteriormente, las plantas se marchitan y también pueden quebrarse en la mitad del tallo debido a que no pueden soportar el peso del capítulo (Figura 10) (Muntañola-Cvetkovic et al. 1981).
Se cree que el cancro del tallo por Phomopsis tiene solo un ciclo de enfermedad primario que involucra ascosporas que se desplazan por distancias considerables desde los reservorios de inóculo en los rastrojos, malezas hospedantes u otros campos de girasoles infestados. Las ascosporas infectan las plantas de girasol en el campo principalmente debido a la falta de movimiento del aire y la mayor humedad relativa en lotes sembrados con alta densidad o elevado canopeo (Mihaljcevic et al. 1985).
Manejo de la enfermedad
Labranza: Se sabe que las especies de
Diaporthe sobreviven en rastrojos de cultivos anteriores hasta por cinco años (Masirevic y Gulya 1992). Enterrar los residuos infectados mediante la labranza reducirá drásticamente los niveles de inóculo (Gulya et al. 1997), pero la tasa de descomposición de los residuos de cultivos dependerá de las condiciones climáticas invernales.
Rotación: Una pausa rotacional de dos a cuatro años con cultivos que no sean hospedadores de
Diaporthe (por ejemplo, cultivos de grano fino y maíz (Zea mays L.)) pueden reducir el inóculo (Masirevic y Gulya 1992).
Densidad de plantas y fertilización con nitrógeno: Menores densidades y menores niveles de nitrógeno pueden reducir la incidencia y la gravedad del cancro del tallo por Phomopsis (Debaeke y Moinard 2010). Debaeke y Moinard (2010) realizaron un estudio a campo que abarcó dos ciclos de siembra observando que las parcelas que recibieron fertilización de nitrógeno alta (120 kg N ha-1) tenían un número significativamente mayor de lesiones alrededor de los tallos por planta en comparación con las parcelas sin fertilización. Además, las parcelas con mayor cantidad de plantas (8 plantas m-2 = 80.000 plantas ha-1) tuvieron un mayor número de lesiones de tallo en comparación con las parcelas con una densidad de plantas menor (5 plantas m-2 = 50.000 plantas ha-1) (Debaeke y Moinard 2010). En los Estados Unidos, habitualmente se utilizan densidades de 50.000 plantas ha-1 o menores.
Tratamiento sanitario de semillas: Aunque las especies de
Diaporthe potencialmente pueden ser transmitidas por semillas, hay evidencia limitada acerca de la transmisión de los patógenos del girasol por esta vía (Fayret et al. 1996; Slyusar y Bochkarev 1997; S. Thompson, inédito).
Manejo de malezas: En general, las malezas son reservorios importantes de inóculo de especies de
Diaporthe asociadas con el girasol tanto como puentes vivos (verdes) o muertos (marrones) (Thompson et al. 2015). Entre estas malezas se incluyen,
Helianthus annuus,
Xanthium italicum,
X. strumarium,
X. pungens y
Arctium lappa ya que son hospedadores de
D. helianthi (Mihaljcevic et al. 1985; Vrandecic et al.2010). Entre los hospedadores de
D. gulyae se encuentran la soja (Glycine max L.) y el maíz (Zea mays L.) y entre las malezas la lista abarca
Kochia scoparia,
Chenopodium album,
Polygonum pensylvanicum y
Carthamus lanatus. (Ash et al. 2010; Mathew et al. 2018b; S. Markell y F. Mathew, inédito; Thompson et al. 2015). Entre los hospedadores de
D. gulyae, el maíz,
Kochia scoparia y
Chenopodium album son asintomáticos (S. Markell y F. Mathew, inédito; Thompson et al. 2015). En definitiva, el hecho de que la soja, el maíz y las malezas puedan servir como hospedadores alternativos para
D. helianthi o
D. gulyae contribuyendo a mantener el inóculo, es un motivo relevante para que se le preste especial atención al manejo de estas especies en los lotes que sean destinados al cultivo de girasol.
Control químico: En Europa se han utilizado fungicidas para controlar el cancro del tallo por Phomopsis. Por ejemplo, en un estudio de Debaeke et al. (2003) se observó que la aplicación de un fungicida protector en el estadio fenológico R1 redujo la incidencia de la enfermedad. Además, Debaeke et al. (2003) recomendaron la aspersión de fungicidas a una altura de planta de 50 a 70 cm utilizando equipos pulverizadores convencionales. En los Estados Unidos, aunque los fungicidas están etiquetados para el manejo de las enfermedades foliares del girasol, datos preliminares de S. Markell y R. Harveson (inédito) indicaron que los fungicidas por sí solos no son suficientes para el manejo del cancro del tallo por Phomopsis. Olson (2017) evaluó un enfoque integrado que incluía la combinación de la aplicación de fungicidas con la siembra de híbridos de girasol con tolerancia a la enfermedad. Bajo una severa presión de enfermedades, Olson (2017) observó mayores rendimientos en parcelas con una sola aplicación de piraclostrobina (grupo 11,
Fungicide Resistance Action Committee) en la etapa de inicio de la yema (etapa de crecimiento R1) en híbridos susceptibles y tolerantes en comparación con parcelas control no tratadas. Sin embargo, una vez que aparecen los síntomas de la enfermedad, las aplicaciones de fungicidas pueden no ser efectivas (Olson 2017).
Resistencia genética: Algunos estudios han sugerido que la resistencia del girasol a
D. helianthi es de naturaleza cuantitativa con efectos aditivos (Viguié et al. 1999). Además, Degener et al. (1999) propusieron que los factores de resistencia en la hoja y el tallo se heredan de forma independiente. En la última década, el germoplasma de girasol de recursos
ex situ (accesiones del banco de semillas del USDA) e
in situ (especies silvestres y variedades autóctonas) se evaluó en condiciones de campo para determinar su resistencia al cancro del tallo por Phomopsis. Gulya (1997) evaluó un total de 1106 accesiones de la colección de girasol cultivado del USDA y observó que el 2% del total de accesiones tenía resistencia a la enfermedad (definida como una proporción menor del 10% de plantas infectadas). Además de la colección de girasoles cultivados por el USDA, se informó que varias especies silvestres de
Helianthus tienen resistencia a
D. helianthi (Cuk 1982; Dozet 1990; Škorić 1984). Talukder et al. (2014) han identificado 30 líneas parentales de Rusia y Europa que tienen resistencia a
D. helianthi. Recientemente, Mathew et al. (2018a) seleccionaron 54 accesiones de girasol cultivadas por el USDA para determinar su resistencia a
D. helianthi y
D. gulyae en el invernadero utilizando como control de susceptibilidad la línea endocriada de girasol confitero HA288. Entre las 54 accesiones estudiadas, 13 fueron resistentes a
D. helianthi y 4 a
D. gulyae (Mathew et al. 2018a).
Significado
El cancro del tallo por Phomopsis es una enfermedad fúngica responsable de significativas pérdidas de rendimiento en girasol, tanto en los Estados Unidos como en otros países del mundo. En lotes comerciales de girasol se han observado pérdidas de rendimiento en grano del 30 al 40% como resultado del marchitamiento y el vuelco de las plantas provocados por la infección temprana por
Diaporthe (Figura 10; Masirevic y Gulya 1992; Mathew et al. 2015b; Thompson et al. 2011). En un estudio de Pinochet y Estragnat (1996) se observó que las lesiones alrededor de los tallos pueden causar pérdidas de rendimiento de alrededor de 0,15 tn ha-1 por cada 10 % de plantas con lesión del tallo, independientemente del genotipo y el nivel de resistencia. Además del rendimiento en grano, el contenido de aceite también puede reducirse entre 15 % y 25 % debido a la enfermedad (Aćimović 1986; Debaeke y Moinard 2010). Actualmente, las opciones para manejar el cancro del tallo por Phomopsis son limitadas en los Estados Unidos y otros países productores de girasol del mundo. Por ejemplo, en los Estados Unidos, solo se cuenta con el control químico con fungicidas del grupo 11-FRAC y la siembra de híbridos de girasol parcialmente resistentes (Olson 2017) aunque se están realizando más investigaciones para identificar las mejores estrategias destinadas al manejo eficiente del cancro del tallo de girasol por Phomopsis.
Referencias bibliográficas seleccionadas
Aćimović, M. 1986. The effect of
Phomopsis sp. infection on grain yield and oil content of sunflower plants. Helia 9: 73–76.
Aćimović, M., and Štraser, N. 1982. Phomopsis sp. - novi parazit suncokreta (Phomopsis sp. - new parasite in sunflower). Zaštita Bilja 33(2): 117-158 (English summary).
Androsova, V., Balakhnina, I., and Gulya, T. 2008. Structural aspects regarding formation and emission of
Diaporthe (Phomopsis)
helianthi ascospores. In: 17th International Sunflower Conference Program and Abstracts, Córdoba, Spain. June 8-12, 2008. pp. 103-107.
Ash, G. J., Sakuanrungsirikul, S., Anschaw, E., Stodart, B. J., Crump, N., Hailstones, D., and Harper, J. D. I. 2010. Genetic diversity and phylogeny of a
Phomopsis sp., a putative biocontrol agent for
Carthamus lanatus. Mycologia 102: 54– 61.
Berglund, D. R. (eds.) 2007. Sunflower production. North Dakota State University. Extension Service Report No. 25. Fargo, North Dakota, USA.
Bogdanova, V. N., Karadzova, L.V., and Steinberg, M.E. 1986. Vovremia obnaruzit Phomopsis. Seljskoe hozjaistvo Moldavii 12: 24-25.
Cuk, L. 1982. Using wild species in sunflower breeding. Uljarstvo 1: 23-27.
Debaeke, P., Bedoussac, L., Bonnet, C., Bret-Mestries, E., Seassau, C., Gavaland, A., Raffaillac, D., Tribouillois, H., Véricel, G., and Justes, E. 2017. Sunflower crop: environmental-friendly and agroecological. OCL 24(3): D304. DOI: 10.1051/ocl/2017020.
Debaeke, P., and Moinard, J. 2010. Effect of crop management on epidemics of Phomopsis stem canker (Diaporthe helianthi) for susceptible and tolerant sunflower cultivars. Field Crops Res. 115: 50-60.
Debaeke, P., Estragnat, A., and Reau, R. 2003. Influence of crop management on sunflower stem canker (Diaporthe helianthi). Agronomie, EDP Sciences. 23 (7): 581-592.
Degener, J., Melchinger, E. A., and Hahn, V. 1999. Inheritance of resistance to
Phomopsis in sunflower: Study of leaf and stem resistance after artificial and natural infection. Helia 22(31): 105-115.
Dozet, B. M. 1990. Resistance to
Diaporthe/Phomopsis helianthi Munt. -Cvet. et al. wild sunflower species. In: Proceedings of the 12th Sunflower Research Workshop, Fargo, ND. January 9–10. 1990. National Sunflower Association, Bismarck, North Dakota, USA, pp. 86–88.
Fayret, J., Quenin, H., and Perny, A. 1996. Evolution of sanitary state of sunflower seeds. Consequences of capitulum attacks by
Phomopsis helianthi and
Phoma macdonaldii [French]. Phytoma - La Défense des Végétaux 487: 37-40.
Gulya, T. J., Rashid, K. Y., and Masirevic, S. M. 1997. Sunflower diseases. In: Schneiter, AA (ed), Sunflower technology and production, American Society of Agronomy, Madison, Wisconsin, USA, pp. 313–319.
Gulya, T. J. 1997. Phomopsis stem canker resistance in USDA and commercial sunflower germplasm. Proc. 19th Sunflower Res. Workshop, Fargo, ND, USA, pp. 76-78.
Hajdu, F., Baumer, J. S., and Gulya, T. 1984. Occurrence of Phomopsis stem canker in Minnesota and North Dakota. Proc. Sunflower Research Workshop, Bismarck, North Dakota, p. 15.
Harveson, R. M., Markell, S. G., Block, C. C., and Gulya, T. J. 2016. Compendium of Sunflower Diseases, 1st ed. American Phytopathological Society, St. Paul, MN, USA.
Herr, L. J., Lipps, P. E., and Watters, B. L. 1983. Diaporthe stem canker of sunflower. Plant Dis. 67: 911-913.
Lamarque, C., and Perny, R. A. 1985. Nouvelle maladie du tournesol: Le phomopsis. Cultivar 179: 57-59.
Li, S., Maric, A., and Masirevic, S. 1985. Biological and epidemiological studies of Phomopsis sp. (Diaporthe sp.) in sunflower. Zastita Bilja 36: 357-370.
Maric, A., Masirevic, S., and Li, S. 1982. Prilog proucavanju Phomopsis spp. (Diaporthe spp.) prowsrokovace sive pecevosti stabea suncokreta. Zastita Bilja 33(4): 403-419.
Masirevic, S., and Gulya, T. J. 1992. Sclerotinia and Phomopsis – two devastating sunflower pathogens. Field Crop Res. 30: 271– 300.
Mathew, F., Olson, T., Marek, L., Gulya, T., and Markell, S. 2018a. Identification of sunflower (Helianthus annuus) accessions resistant to Diaporthe helianthi and Diaporthe gulyae. Plant Health Prog. 19: 97–102.
Mathew, F., Gulya, T. J., Jordahl, J., and Markell, S. 2018b. First report of stem disease of soybean (Glycine max) caused by Diaporthe gulyae in North Dakota. Plant Dis. 102: 240.
Mathew, F. M., Rashid, K. Y., Gulya, T. J., and Markell, S. G. 2015a. First report of Phomopsis stem canker of sunflower (Helianthus annuus) caused by Diaporthe gulyae in Canada. Plant Dis. 99: 160.
Mathew, F. M., Alananbeh, K. M., Jordahl, J. G., Meyer, S. M., Castlebury, L. A., Gulya, T. J., and Markell, S. G. 2015b. Phomopsis stem canker: A reemerging threat to sunflower (Helianthus annuus) in the United States. Phytopathology 105: 990-997.
Mathew, F. M. 2014. Molecular characterization and pathogenicity of sunflower stem pathogens (Doctoral dissertation). Retrieved from ProQuest Dissertations Publishing Database. (Accession No. 3629478) (Last Accessed: 11/28/2017).
Mihaljcevic, M., Muntanola-Cvetkovic, M., Vukojevic, J., and Petrov, M. 1985. Source of infection of sunflower plants by Diaporthe helianthi in Yugoslavia. Phytopathol. Z. 113: 334-342.
Mihailova, P. 1984. Savremeni fitopatogeni problemi pri slanogleda. Rastitelna zascita 4(84): 11-14.
Miric, E. 2002. Pathological, morphological and molecular studies of a worldwide collection of the sunflower pathogens Phomopsis helianthi and Phoma macdonaldii. PhD thesis, University of Queensland, Australia.
Muntañola-Cvetkovic, M., Vukojevic, J., and Mihaljčević, M. 1991. The systemic nature of the sunflower disease caused by Diaporthe helianthi. Can. J. Bot. 69: 1552-1556.
Muntañola-Cvetkovic, M., Vukojevic, J., and Mihaljčević, M. 1989. Pathohistology of sunflower stems attacked by Diaporthe helianthi. Can. J. Bot. 67: 1119-1125.
Muntañola-Cvetkovic´, M., Mihaljčević, M., and Petrov, M. 1985. On the identity of the causative agent of a serious Phomopsis-Diaporthe disease in sunflower plants. Nova Hedwigia 34: 417–435.
Nemeth, F., Princzinger, G., and Vörös, J. 1981. New disease in Hungary. Magyar Mezogazdasag 48: 10-11.
Olson, T. R., Kontz, B., Gulya, T. J., Markell, S. G., and Mathew, F. M. 2017. First report of Diaporthe stewartii causing Phomopsis stem canker of sunflower (Helianthus annuus) in Minnesota. Plant Dis. 101: 382.
Olson, T. R. 2017. Managing Phomopsis stem canker of sunflower using improved diagnosis and quantification of the causal pathogens. Theses and Dissertations. 1184. http://openprairie.sdstate.edu/etd/1184
Pericin, D., Jarak, M., Anotv, M., and Dozet, B. 1994. Pectinase from Phomopsis helianthi-the agent of sunflower stem canker. Helia 17(20): 21-30.
Pinochet, X., and Estragnat, A. 1996. Field evaluation of hybrid sensitivity to Diaporthe helianthi: relationship between symptoms and yield losses. In: Proceedings of the 14th International Sunflower Conference, Beijing, China, pp. 777–780.
Pinochet, X. 1995. Phomopsis et evaluation varie´tale de la sensibilite´. In: Le Page, R. (Ed.), Le Phomopsis du tournesol. CETIOM, Paris, France, pp. 27–36.
Regnault, Y. 1985. Premieres observations sur le Phomopsis du tournesol. Bullet Inf. Techn. CETIOM, Paris, France. 92: 13-20.
Santos, J. M., Correia, V. G., and Phillips, A. J. 2010. Primers for mating-type diagnosis in Diaporthe and Phomopsis: their use in teleomorph induction in vitro and biological species definition. Fungal Biol. 114: 255–270.
Scripka, O. V., Sheluhin, V. I., Petina, V. V., and Serebryakova, P. P. 1993. Phomopsis in sunflower. Crop Prot. 8: 24-25.
Škorić, D. 1984. Genetic resources in the Helianthus genus. In: D. Škorić, editor, Proceedings of the International Symposium on Science and Biotechnology for an Integral Sunflower Utilization, Bari, Italy. October 25, 1984. pp. 37–73.
Slyusar, E. L., and Bochkarev, N. I. 1997. Phomopsis and seed quality. Zashchita Karantin Rastenić 8: 30–31.
Su, L., Maric, A., and Masirevic, S. 1985. Biological and epidemiological studies of
Phomopsis sp. (Diaporthe sp.) in sunflower. Zaštita Bilja 36: 357-370.
Talukder, Z. I., Hulke, B. S., Marek, L. F., and Gulya, T. J. 2014. Sources of resistance to sunflower diseases in a global collection of domesticated USDA Plant Introductions. Crop Sci. 54: 694-705.
Thompson, S. M., Tan, Y. P., Neate, S. M., Grams, R. M., Shivas, R. G., Lindbeck, K., and Aitken, E. A. B. 2018. Diaporthe novem isolated from sunflower (Helianthus annuus) and other crop and weed hosts in Australia. Eur. J. Plant Pathol. 91: 1-9.
Thompson, S.M., Tan, Y.P., Shivas, R.G., Neate, S.M., Morin, L, Bissett, A., and Aitken, E.A.B. 2015. Green and brown bridges between weeds and crops reveal novel Diaporthe species in Australia. Persoonia 35: 39-49.
Thompson, S. M., Tan, Y. P., Young, A. J., Neate, S. M., Aitken, E. A. B., and Shivas, R. G. 2011. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia 27: 80–89.
Viguié, A., Vear, F., and de Labrouhe, D. T. 1999. Interactions between French isolates of Phomopsis/Diaporthe helianthi Munt. -Cvet. et al. and sunflower (Helianthus annuus L.) genotypes. Eur. J. Plant Pathol. 105: 693-702.
Vrandecic, K., Jurkovic, D., Riccioni, L., Cosic, J., and Duvnjak, T. 2010. Xanthium italicum, Xanthium strumarium, and Arctium lappa as new hosts for Diaporthe helianthi. Mycopathologia 170: 51–60.
Yakutkin, V. I. 1988. Methods for identification and evaluation of sunflower Phomopsis. In: Collection of methodological recommendations in crop protection. St. Petersburg, Russia, pp. 191-207.
Yang, S. M., and Gulya, T. J. 1984. Groups of Diaporthe/Phomopsis isolates obtained from cultivated sunflower. Phytopathology 74: 868.
Yang, S. M., Berry, R. W., Luttrell, L. E. S., and Vongkaysone, T. 1984. A new sunflower disease in Texas caused by
Diaporthe helianthi. Plant Dis. 68: 254-255.
Zambelli A., Mancebo, M. F., Bazzalo, M. E., Reid, R. J., Sanchez, M. C., Kontz, B. J., and Mathew, F. M. 2021. Six species of Diaporthe associated with Phomopsis stem canker of sunflower in southern Pampean region of Argentina. Plant Health Prog. 22: 136-142.

